Inflammasome dysregulation is associated with various inflammatory diseases. This review compares the mechanisms of canonical NLRP3 inflammasome-mediated Caspase-1 activation and the non-canonical pathway in which Caspase-11 (human Caspase-4/5) directly senses intracellular LPS for independent activation. Using IL-1β/IL-18 maturation and GSDMD cleavage as functional readouts, we elucidate the differences between the two pathways in substrate processing and pyroptosis induction. Furthermore, we summarize the progress of small-molecule inhibitors targeting NLRP3 (e.g., MCC950, CY-09) and GSDMD (e.g., disulfiram), and discuss the therapeutic potential and clinical translation obstacles of targeting pyroptosis pathways. Through integrated analysis, this review aims to provide a reference for understanding inflammasome-mediated immune regulation and precision intervention strategies.
Table of Contents
1. Canonical NLRP3 inflammasome activation leading to Caspase-1 cleavage
2. Non-canonical inflammasome pathway mediated by Caspase-11 in mice
3. LPS intracellular sensing and Caspase-11 activation mechanism
4. IL-1β and IL-18 maturation as functional readouts of Caspase-1 activity
5. Macrophage pyroptosis induced by Caspase-1 vs Caspase-11
6. Therapeutic targeting of inflammasome-driven pyroptosis pathways
01 Canonical NLRP3 inflammasome activation leading to Caspase-1 cleavage
The NLRP3 inflammasome is a multiprotein complex composed of the sensor protein NLRP3 (containing an N-terminal PYD domain, a central NACHT domain, and C-terminal LRR domains), the adaptor protein ASC (containing PYD and CARD domains), and the effector protein pro-caspase-1. In the resting state, NLRP3 adopts an ADP-bound self-inhibited conformation. Caspase-1 (cysteine-aspartic protease 1), an inflammatory caspase, was originally discovered in 1989 and named IL-1β-converting enzyme[1]. It typically exists in the cytosol as an inactive zymogen and is activated within various inflammasome complexes, such as those assembled by NLRP3, NLRC4, and AIM2[2].
Canonical NLRP3 inflammasome activation follows a typical two-signal mechanism. Signal 1 (Priming): PAMPs such as LPS activate the NF-κB pathway via TLR4, upregulating the transcriptional expression of NLRP3 and pro-IL-1β. In addition, priming also exerts non-transcriptional effects, through the MyD88-IRAK4-IRAK1 signaling axis, it mediates post-translational modifications such as deubiquitination of NLRP3, thereby rendering NLRP3 in a “primed” state. Signal 2 (Activation): Various stimuli (extracellular ATP, nigericin, monosodium urate crystals, cholesterol crystals, silica, etc.) trigger NLRP3 activation through a common converging mechanism: potassium efflux (K⁺ efflux). Other converging signals include lysosomal damage, mitochondrial reactive oxygen species (mtROS) production, and release of oxidized mitochondrial DNA[3].
Sensing of activation signals induces conformational changes in NLRP3, leading to ADP release and exposure of the oligomerization interface. Key steps include:
The permissive role of NEK7: NEK7 (NIMA-related kinase 7) is an essential co-activator for NLRP3 activation. Cryo-electron microscopy structures have shown that NEK7 binds to NLRP3 through its HD2 and LRR domains, forming a "bridge" between adjacent NLRP3 molecules to facilitate oligomerization assembly. The kinase activity of NEK7 is not required for NLRP3 activation; its role is primarily structural[3].
ASC speck formation: Activated NLRP3 recruits ASC through PYD-PYD homotypic interactions, and ASC further recruits pro-caspase-1 via CARD-CARD interactions, forming large (~1 μm) perinuclear ASC speck structures.
Proximity-induced autoactivation of caspase-1: On the inflammasome platform, the high local concentration of pro-caspase-1 promotes homodimerization and autocleavage, generating the active heterotetrameric caspase-1 composed of p20 and p10 subunits. Upon activation, caspase-1 exerts multiple functions, including: ① cleavage of pro-IL-1β and pro-IL-18 into bioactive pro-inflammatory cytokines; ② cleavage of gasdermin D (GSDMD), releasing its N-terminal domain to form pores in the plasma membrane, thereby inducing a pro-inflammatory programmed cell death termed pyroptosis. In addition, caspase-1 inflammasome is also involved in atypical protein secretion, NF-κB pathway activation, metabolic regulation, and other functions[2,4].
02 Non-canonical inflammasome pathway mediated by Caspase-11 in mice
Caspase-11 is a mouse-specific inflammatory caspase consisting of 373 amino acids, containing an N-terminal CARD domain, as well as p20 and p10 catalytic domains. Its human homologs are caspase-4 and caspase-5. Caspase-11 shares approximately 46% overall amino acid sequence identity with caspase-1, but exhibits significant differences in the substrate-binding pockets (P2–P4)[5].
The activation mechanism of caspase-11 is fundamentally different from that of the canonical inflammasome. Caspase-11 functions as a direct intracellular receptor for LPS. Shi et al. demonstrated that the CARD domain of caspase-11 directly binds the lipid A moiety of LPS (requiring penta-acylated or hexa-acylated lipid A, whereas tetra-acylated lipid A fails to activate). LPS binding induces caspase-11 oligomerization and proximity-induced autoproteolytic activation, with the critical cleavage site at Asp285. Unlike caspase-1, which requires an upstream sensor complex, caspase-11 itself serves as both sensor and effector enzyme: it acts as a pattern recognition receptor for intracellular LPS and also as the protease that executes pyroptosis[6, 7].
Activated caspase-11 mediates the non-canonical inflammasome pathway:
(1)Direct cleavage of GSDMD: Caspase-11 directly cleaves GSDMD at Asp276, releasing the GSDMD-NT pore-forming domain, which forms plasma membrane pores and induces pyroptosis. This effect is independent of canonical inflammasome components, as NLRP3, ASC, and caspase-1 are all dispensable for caspase-11-mediated pyroptosis[8].
(2)Activation of the canonical NLRP3 inflammasome via K⁺ efflux: The pores formed by caspase-11-cleaved GSDMD lead to K⁺ efflux, which is a critical upstream signal for NLRP3 activation[9]. Thus, in the non-canonical pathway, caspase-11 indirectly activates the NLRP3/caspase-1 axis through GSDMD pore-mediated K⁺ efflux, thereby promoting the maturation and release of IL-1β and IL-18[8].
(3)No direct cleavage of pro-IL-1β/pro-IL-18: Caspase-11 itself lacks the ability to cleave pro-IL-1β and pro-IL-18; the maturation of these cytokines relies entirely on caspase-1, which is indirectly activated by caspase-11[5].
In summary, the canonical NLRP3 inflammasome pathway follows a "two-signal" model (priming + activation), mediating IL-1β/IL-18 maturation and GSDMD-dependent pyroptosis through the NLRP3–ASC–caspase-1 axis. In contrast, the non-canonical inflammasome pathway is centered on caspase-11, which serves as a direct intracellular sensor of LPS and is activated independently of the NLRP3/ASC complex. It induces pyroptosis via direct GSDMD cleavage, and further indirectly activates the canonical NLRP3 inflammasome through GSDMD pore-mediated K⁺ efflux, thereby forming an integrated immune response network against Gram-negative bacteria.
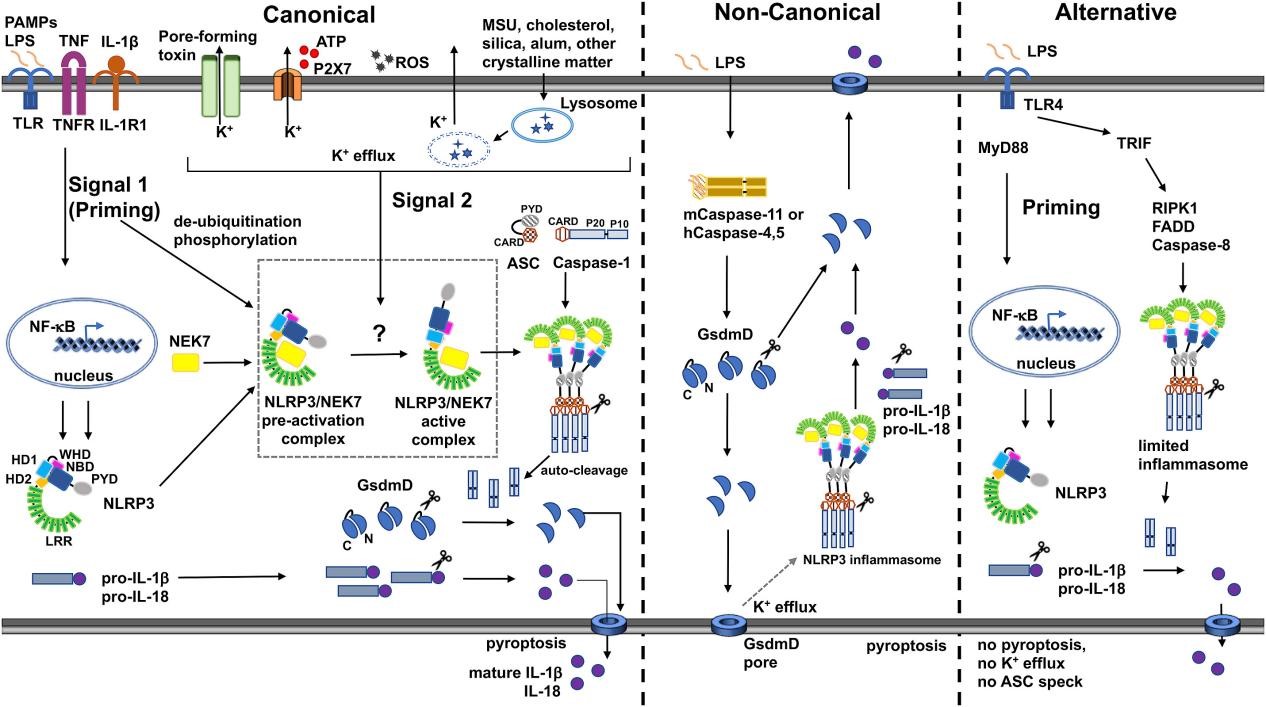
Fig. 1 Canonical, non-canonical, and alternative modes of NLRP3 inflammasome signaling. NLRP3 activation requires two signals: NF-κB priming and PAMP/DAMP stimulation, primarily triggering K⁺ efflux via pore-forming agents, ATP-P2X7, or lysosomal damage. Mitochondrial ROS and oxidized mtDNA also activate NLRP3 via both K⁺-dependent and independent routes. Non-canonical activation by cytosolic LPS through caspase-4/5/11 forms GSDMD pores, causing K⁺ efflux that activates NLRP3. TLR4 in monocytes can activate NLRP3 via RIP1-FADD-caspase-8. The NLRP3-ASC-pro-caspase-1 complex induces caspase-1 auto-cleavage, which processes cytokines and cleaves GSDMD; its N-terminal fragment pores release cytokines and drive pyroptosis[3].
03 LPS intracellular sensing and Caspase-11 activation mechanism
The innate immune system senses pathogen invasion through pattern recognition receptors. LPS from Gram-negative bacteria is one of the most well-studied pathogen-associated molecular patterns (PAMPs). Classical LPS sensing relies on the membrane-bound receptor TLR4; however, a major discovery over the past decade has revealed an intracellular sensing mechanism for LPS-inflammatory caspases-4/5/11 can directly bind cytosolic LPS to trigger the non-canonical inflammasome pathway[5, 6].
The groundbreaking study by Shi et al. published in Nature elucidated the molecular details of direct LPS binding by caspase-11/4/5. Using surface plasmon resonance (SPR) experiments, the researchers found that the CARD domain of caspase-11 directly binds the lipid A moiety of LPS with very high affinity. This binding induces oligomerization of caspase-11, which in turn triggers autoproteolytic activation through a proximity-induced mechanism. Critically, point mutations in the CARD domain (binding-deficient mutants) completely abolished caspase-11 responsiveness to LPS, confirming that the CARD domain is indeed the LPS-binding site. Human caspase-4/5 are functionally interchangeable with mouse caspase-11, expression of human caspase-4/5 in caspase-11-deficient mouse macrophages fully restored responsiveness to cytosolic LPS[10].
The primary substrate of activated caspase-11 (as well as caspase-1 in the canonical pathway) is GSDMD. GSDMD consists of an N-terminal pore-forming domain (PFD) and a C-terminal repressor domain (RD) connected by a linker. Caspase-11/1 cleave GSDMD at the linker at mouse Asp276 (human Asp275), releasing an approximately 31 kDa N-terminal fragment (GSDMD-NT). GSDMD-NT then translocates to the inner leaflet of the plasma membrane, binds to negatively charged phospholipids such as phosphatidylinositol-4,5-bisphosphate (PIP2) and cardiolipin, and oligomerizes into ring-shaped structures, forming transmembrane pores with an inner diameter of approximately 10-18 nm[11,12]. These pores not only disrupt the barrier function of the plasma membrane, leading to loss of ion gradients and water influx, but more importantly, they provide physical channels for the release of cellular contents.
%20(E-CK-A484)%20and%20analyzed%20by%20flow%20cytometry_.jpg)
Fig. 2 Jurkat cells were stained with Caspase 4 Substrates (Green) (E-CK-A484) and analyzed by flow cytometry (The data are provided by Elabscience).
04 IL-1β and IL-18 maturation as functional readouts of Caspase-1 activity
The most classic and best-characterized function of active caspase-1 is the cleavage of pro-IL-1β (31 kDa) and pro-IL-18 (24 kDa) to generate their mature active forms (17 kDa and 18 kDa, respectively). Caspase-1 cleaves pro-IL-1β at Asp116 (human), releasing biologically active mature IL-1β. This processing is highly specific; although other proteases such as caspase-8 can also cleave pro-IL-1β under certain conditions, caspase-1 is the primary processing enzyme under physiological conditions. Both IL-1β and IL-18 lack classical signal peptide sequences and therefore cannot be released through the classical ER-Golgi secretory pathway. They are secreted via "unconventional secretion," a process that is tightly coupled to caspase-1 activation and the cell death program[4].
In the non-canonical inflammasome pathway, the maturation of IL-1β and IL-18 follows a "two-step" mechanism: First, cytosolic LPS activates caspase-11, which cleaves GSDMD to form pores, leading to K⁺ efflux and indirect activation of the NLRP3 inflammasome; Second, the NLRP3 inflammasome activates caspase-1, which directly cleaves pro-IL-1β and pro-IL-18. Therefore, detection of mature IL-1β and IL-18 serves as a functional readout of caspase-1 activity, while also reflecting the degree of canonical inflammasome engagement in the non-canonical pathway. Studies have shown that in caspase-11-deficient macrophages infected with Gram-negative bacteria, although pyroptosis is markedly reduced, the levels of mature IL-1β and IL-18 are also significantly diminished, confirming the importance of this cascade.
Detection of mature IL-1β and IL-18 has become the gold-standard functional readout for evaluating caspase-1 activity. In in vitro studies, the level of mature IL-1β (p17) in cell supernatants, measured by ELISA or immunoblotting, can quantitatively reflect the degree of caspase-1 activation. In animal models and clinical samples of inflammasome-associated diseases, such as cryopyrin-associated periodic syndromes (CAPS), familial Mediterranean fever (FMF), and gout, circulating levels of IL-1β and IL-18 serve as important biomarkers for disease activity and therapeutic response. In recent years, IL-1β/IL-18 release assays have also been widely employed in drug screening: by measuring the inhibitory effects of compounds on IL-1β maturation and release following inflammasome activation, their potential for targeting caspase-1/NLRP3 can be evaluated[13].
%20in%20Raw264_7%20cells%20after%20pyroptosis_.jpg)
Fig. 3 Detection of caspase-1 enzymatic activity (E-CK-A381) in Raw264.7 cells after pyroptosis. Raw264.7 cells were co-stimulated with LPS and ATP to induce pyroptosis, then incubated with a caspase-1 probe and imaged under a fluorescence microscope. In parallel, the results were analyzed using a caspase-1 absorbance-based activity assay kit. The results showed that caspase-1 enzymatic activity was significantly increased in Raw264.7 cells after pyroptosis (The data are provided by Elabscience).
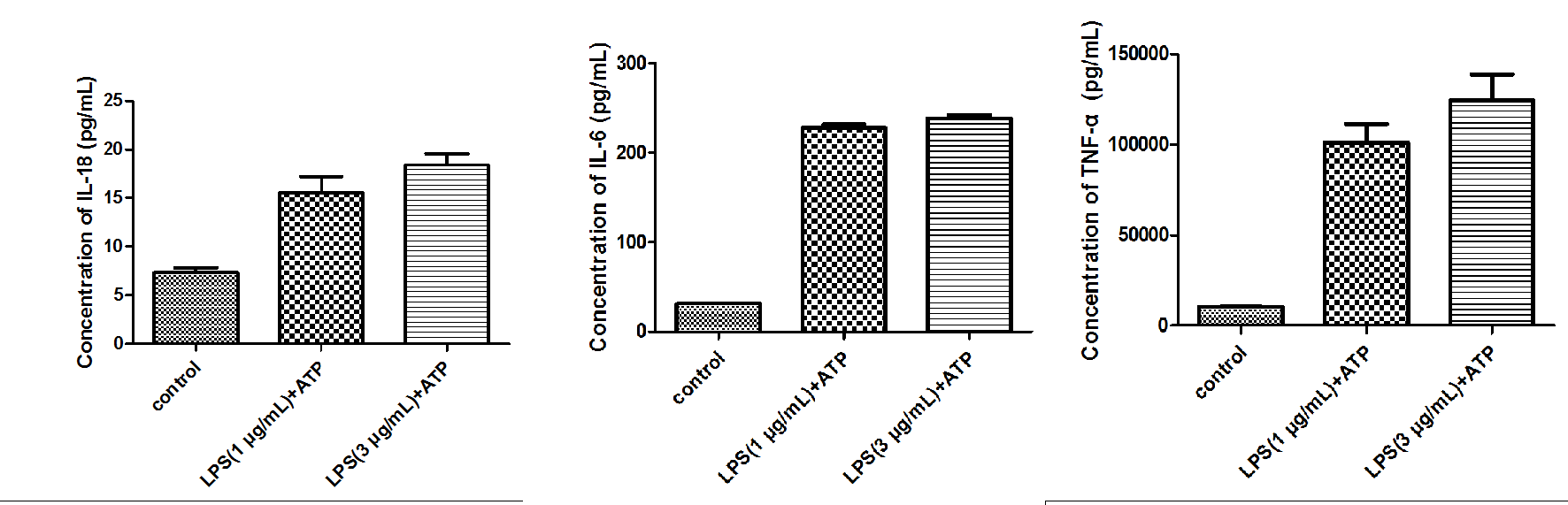
Fig. 4 Detection of inflammatory cytokines in Raw264.7 cells after pyroptosis. Raw264.7 cells were co-stimulated with LPS and ATP to induce pyroptosis. The cell supernatants were then collected, and the levels of secreted cytokines were measured using mouse TNF-α, IL-6, and IL-18 elisa kits (E-EL-M0730, E-EL-M0044, E-EL-M3063). The results showed that the secretion and release of IL-18, IL-6, and TNF-α in the supernatants of pyroptotic cells were significantly increased (The data are provided by Elabscience).
05 Macrophage pyroptosis induced by Caspase-1 vs Caspase-11
Pyroptosis is a pro-inflammatory programmed cell death characterized by cell swelling, plasma membrane pore formation, and release of pro-inflammatory cytokines (IL-1β, IL-18). The canonical inflammasome pathway mediates pyroptosis through activation of Caspase-1, whereas the non-canonical pathway is directly driven by Caspase-11 sensing intracellular LPS. Although both pathways rely on GSDMD pore formation for pyroptosis execution, they exhibit significant differences in activation mechanisms, substrate selectivity, and biological functions.
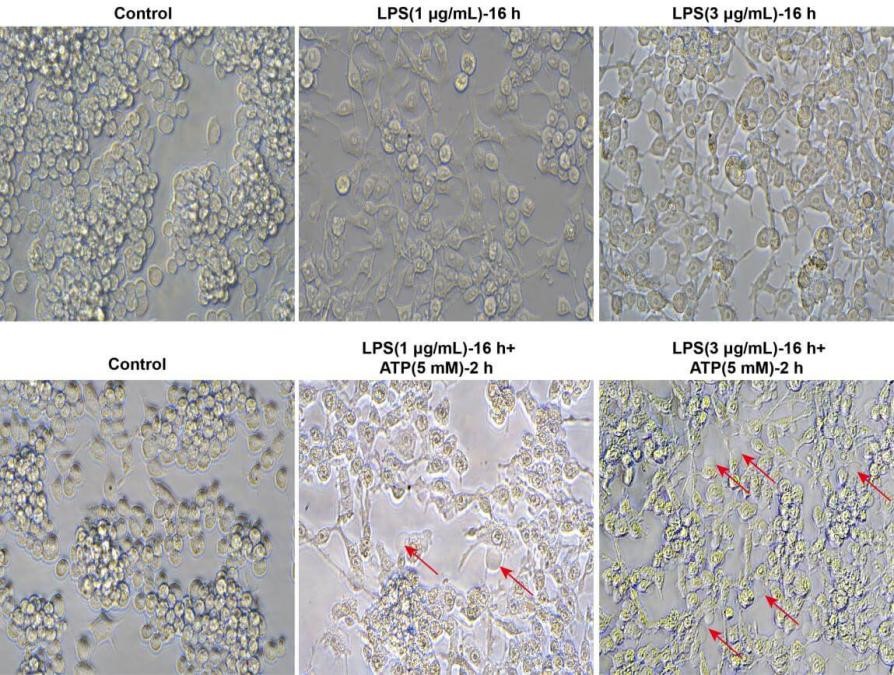
Fig. 5 Morphological observation of pyroptosis in Raw264.7 cells. As shown in the figure, Raw264.7 cells were stimulated with LPS alone for 16 h, and microscopic observation revealed increased granularity within the cells, but no obvious balloon-like blebbing characteristic of pyroptosis. After co-stimulation with LPS plus ATP (5 mM) for 2 h, the typical blebbing of pyroptotic cells (indicated by red arrows) was clearly observed under the microscope. (Typical morphological features of pyroptosis include: cell swelling and deformation, distorted organelles, formation of pyroptotic bodies, balloon-like blebbing, and rupture of the plasma membrane) (The data are provided by Elabscience).
5.1 The differences in activation mechanisms
Canonical inflammasome pathway: Caspase-1 activation depends on the inflammasome platform. Sensor proteins (such as NLRP3, NLRC4, and AIM2) recognize PAMPs/DAMPs and recruit ASC through homotypic interactions. ASC then recruits pro-caspase-1, which undergoes proximity-induced dimerization and autocleavage on the platform to generate the active enzyme[14].
Non-canonical inflammasome pathway: The CARD domain of caspase-11 directly binds the lipid A moiety of intracellular LPS, requiring no upstream sensors or adaptor proteins to induce its oligomerization and autoproteolytic activation. This process also requires GBPs to help disrupt the bacterial outer membrane or pathogen-containing vacuoles, thereby exposing the sequestered lipid A for caspase-11 recognition[6,14,15].
5.2 The difference in substrate selectivity
Processing of IL-1β and IL-18: Caspase-1 efficiently cleaves pro-IL-1β and pro-IL-18, with kcat/KM values reaching >10⁵ M⁻¹s⁻¹, whereas caspase-11 cleaves these cytokine precursors with extremely low efficiency (kcat/KM ≈ 9 M⁻¹s⁻¹), which is more than 10,000-fold lower than that of caspase-1. It has been estimated that at physiological concentrations, caspase-11 would require over 3 months to cleave 50% of pro-IL-1β, whereas caspase-1 needs only 5–10 minutes.
Cleavage of GSDMD: Both enzymes efficiently cleave GSDMD to generate the pore-forming N-terminal domain, with caspase-1 being only approximately 13-fold more efficient than caspase-11.
Differences in substrate-binding pockets: Structural analyses reveal significant differences in the P2–P4 substrate-binding pockets between caspase-1 and caspase-11. Caspase-1 prefers tryptophan at the P4 position, whereas caspase-11 prefers aliphatic amino acids such as valine[16].
5.3 Indirect pathways for cytokine maturation
Although Caspase-11 cannot directly process IL-1β and IL-18, it promotes their maturation through indirect mechanisms. The plasma membrane pores formed by Caspase-11–cleaved GSDMD lead to K⁺ efflux, a signal sensed by the NLRP3 inflammasome, which subsequently activates Caspase-1. Caspase-1 then processes pro-IL-1β and pro-IL-18. Therefore, the release of IL-1β in the non-canonical pathway depends on the dual action of GSDMD pores and the NLRP3–Caspase-1 axis.
5.4 The differences in non-pyroptosis effect functions
Recent studies have revealed that Caspase-11 possesses multiple biological functions independent of pyroptosis. A research systematically summarizes the non-pyroptotic effects of Caspase-11[17]:
Regulation of actin polymerization: Caspase-11 regulates the phosphorylation status of cofilin via RhoA and Slingshot proteins, thereby modulating actin dynamics and promoting the fusion of pathogen-containing phagosomes with lysosomes.
Promotion of autophagy: Caspase-11 positively regulates autophagosome formation and their trafficking to lysosomes. In macrophages infected with Burkholderia cenocepacia, loss of Caspase-11 leads to impaired autophagosome maturation.
Direct cleavage of IL-1α: Studies have shown that Caspase-11 and its human homolog Caspase-5 can directly cleave pro-IL-1α to generate a more active mature form. This function is of particular significance in the senescence-associated secretory phenotype (SASP).
Regulation of neutrophil extracellular trap (NET) formation: In pulmonary infections, Caspase-11 promotes the release of NETs through a GSDMD-dependent pathway, thereby contributing to innate immune defense[15].
5.5 Sensing differences in pathogens
Caspase-1 senses a variety of PAMPs/DAMPs (such as ATP, flagellin, double-stranded DNA, etc.) through upstream sensors, whereas Caspase-11 specifically recognizes intracellular LPS with stringent gating for the acylation state of lipid A, only the hexa-acylated form is effective, while penta- and tetra-acylated forms are not. As a result, certain pathogens can evade recognition by modifying their LPS structure. Furthermore, Caspase-1 depends on TLR4 to provide a priming signal, whereas the transcription of Caspase-11 is induced by the TLR4–TRIF–IFN–STAT1 axis, with very low expression in resting cells that requires LPS or IFN-γ stimulation for upregulation. In contrast, human Caspase-4 is constitutively expressed, suggesting species-specific differences in immune surveillance strategies[15,17].
%20in%20Raw264_7%20cells%20after%20pyroptosis_.jpg)
Fig. 6 Flow cytometric detection of ROS (E-BC-K138-F) in Raw264.7 cells after pyroptosis. Raw264.7 cells were induced to undergo pyroptosis by co-stimulation with LPS and ATP, and then ROS levels were detected using a ROS probe and analyzed by flow cytometry. As shown in the figure, ROS accumulation was significantly increased in pyroptotic cells (The data are provided by Elabscience).
06 Therapeutic targeting of inflammasome-driven pyroptosis pathways
Given the pathogenic roles of aberrant inflammasome activation and pyroptosis in various diseases, intervention strategies targeting this pathway have become an important direction in drug development. Currently, these intervention strategies can be classified according to the location of their targets into the following categories:
6.1 Inhibitors targeting the NLRP3 inflammasome
Aberrant activation of the NLRP3 inflammasome is associated with a variety of diseases, including CAPS (cryopyrin-associated periodic syndromes), Alzheimer’s disease, type 2 diabetes, and gout. The representative inhibitor MCC950 blocks oligomerization by binding to the NACHT domain and has shown efficacy in multiple models; however, its development was terminated in a phase II clinical trial for rheumatoid arthritis due to hepatotoxicity[18-20]. CY-09 similarly targets NACHT to inhibit ATPase activity and orally improves metabolism in diabetic mice. OLT1177 (Dapansutrile) is orally effective and has completed Phase I and entered Phase II trials for gouty arthritis. Tranilast, an approved anti-allergic drug, has been repurposed for inflammatory diseases by blocking NLRP3 assembly. In addition, natural products (such as curcumin analogs, apigenin, etc.) also exhibit inhibitory activity, but their poor water solubility and pharmacokinetic limitations hinder clinical translation.
6.2 Inhibitors targeting Caspase-1
Pralnacasan (VX-740) and VX-765 are early-developed selective inhibitors of Caspase-1. Pralnacasan effectively ameliorated symptoms in the DSS-induced colitis model but was hindered in clinical trials due to hepatotoxicity. Ac-YVAD-CMK is a classical peptide inhibitor of Caspase-1 and has shown protective effects in animal models of Parkinson’s disease[18].
6.3 Strategies targeting GSDMD
GSDMD is a common downstream effector molecule shared by both the canonical and non-canonical pyroptosis pathways, and targeting GSDMD can simultaneously block both pathways. Among representative inhibitors, disulfiram (an FDA-approved drug for alcohol aversion) specifically blocks pore assembly by covalently modifying human GSDMD at Cys191 (mouse Cys192), without affecting GSDMD cleavage or IL-1β processing. It improves survival in sepsis models, with an IC₅₀ of 0.30 μM[21]. Necrosulfonamide (NSA) shares the same Cys-binding site and inhibits p30-GSDMD oligomerization, showing efficacy in septic models as well. Ac-FLTD-CMK, a peptidic inhibitor, acts on the catalytic center of inflammatory caspases and blocks GSDMD cleavage-dependent activation. LDC7559, originally discovered in a phenotypic screen for NETosis inhibitors, has been validated to target GSDMD, although its precise mode of direct interaction is not yet completely elucidated[22].
6.4 Strategies targeting ASC oligomerization and pyroptosis effect molecules
The NBC series (boron-containing compounds): Compounds such as NBC6, developed based on the scaffold of 2-APB (a calcium homeostasis inhibitor), can effectively block ASC speck formation, thereby inhibiting NLRP3 inflammasome assembly, and have shown efficacy in peritonitis models[18].
Quick Overview of Popular Products:
Table 1. Reagents for caspase activity assay and inflammatory factors
|
Cat. No. |
Product Name |
|
E-CK-A381 |
caspase 1 activity assay Kit(Colorimetric Method) |
|
E-CK-A383 |
Caspase 3/7 Activity Assay Kit(Colorimetric Method) |
|
E-CK-A388 |
Caspase 8 Activity Assay Kit(Colorimetric Method) |
|
E-CK-A389 |
Caspase 9 Activity Assay Kit(Colorimetric Method) |
|
E-CK-A382 |
Caspase 2 Activity Assay Kit(Colorimetric Method) |
|
E-CK-A384 |
Caspase 4 Activity Assay Kit(Colorimetric Method) |
|
E-CK-A386 |
Caspase 6 Activity Assay Kit(Colorimetric Method) |
|
E-CK-A481 |
Caspase 1 Activity Detection Substrate for Flow Cytometry |
|
E-CK-A483 |
Caspase 3/7 Activity Detection Substrate for Flow Cytometry |
|
E-CK-A484 |
Caspase 4 Activity Detection Substrate for Flow Cytometry |
|
E-CK-A486 |
Caspase 6 Activity Detection Substrate for Flow Cytometry |
|
E-CK-A488 |
Caspase 8 Activity Detection Substrate for Flow Cytometry |
|
E-CK-A831 |
Caspase 3/7 and Annexin V Double Staining Apoptosis Kit |
|
E-CK-A832 |
Caspase 3/7 and PI Double Staining Apoptosis Kit |
|
E-CK-A833 |
Caspase 3/7 and DAPI Double Staining Apoptosis Kit |
|
E-EL-M0730 |
Mouse IL-18 elisa Kit |
|
E-EL-M0044 |
Mouse IL-6(Interleukin 6) ELISA Kit |
|
E-EL-M3063 |
Mouse TNF-α(Tumor Necrosis Factor Alpha) ELISA Kit |
|
E-EL-M0037 |
Mouse IL-1β elisa Kit |
References:
[1] ZHEN H M, HU Y M, LIU X Y, et al. The protease caspase-1: Activation pathways and functions [J]. BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS, 2024, 717.
[2] SUN Q, SCOTT M J. Caspase-1 as a multifunctional inflammatory mediator: noncytokine maturation roles [J]. JOURNAL OF LEUKOCYTE BIOLOGY, 2016, 100(5): 961-7.
[3] WANG L, HAUENSTEIN A V. The NLRP3 inflammasome: Mechanism of action, role in disease and therapies [J]. MOLECULAR ASPECTS OF MEDICINE, 2020, 76.
[4] SOLLBERGER G, STRITTMATTER G E, GARSTKIEWICZ M, et al. Caspase-I: The inflammasome and beyond [J]. INNATE IMMUNITY, 2014, 20(2): 115-25.
[5] BROZ P, MONACK D M. Noncanonical Inflammasomes: Caspase-11 Activation and Effector Mechanisms [J]. PLOS PATHOGENS, 2013, 9(2).
[6] HUANG X L, FENG Y, XIONG G Q, et al. Caspase-11, a specific sensor for intracellular lipopolysaccharide recognition, mediates the non-canonical inflammatory pathway of pyroptosis [J]. CELL AND BIOSCIENCE, 2019, 9.
[7] AGNEW A, NULTY C, CREAGH E M. Regulation, Activation and Function of Caspase-11 during Health and Disease [J]. INTERNATIONAL JOURNAL OF MOLECULAR SCIENCES, 2021, 22(4).
[8] YI Y S. Functional crosstalk between non-canonical caspase-11 and canonical NLRP3 inflammasomes during infection-mediated inflammation [J]. IMMUNOLOGY, 2020, 159(2): 142-55.
[9] VIGANò E, MORTELLARO A. Caspase-11: The driving factor for noncanonical inflammasomes [J]. EUROPEAN JOURNAL OF IMMUNOLOGY, 2013, 43(9): 2240-5.
[10] SHI J J, ZHAO Y, WANG Y P, et al. Inflammatory caspases are innate immune receptors for intracellular LPS [J]. NATURE, 2014, 514(7521): 187-+.
[11] KAYAGAKI N, STOWE I B, LEE B L, et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling [J]. NATURE, 2015, 526(7575): 666-71.
[12] DING J J, WANG K, LIU W, et al. Pore-forming activity and structural autoinhibition of the gasdermin family [J]. NATURE, 2016, 535(7610): 111-+.
[13] BURDETTE B E, ESPARZA A N, ZHU H, WANG S Z. Gasdermin D in pyroptosis [J]. ACTA PHARMACEUTICA SINICA B, 2021, 11(9): 2768-82.
[14] ROSS C, CHAN A H, VON PEIN J B, et al. Inflammatory Caspases: Toward a Unified Model for Caspase Activation by Inflammasomes [J]. ANNUAL REVIEW OF IMMUNOLOGY, 2022, 40: 249-69.
[15] OH C, VERMA A, AACHOUI Y. Caspase-11 Non-canonical Inflammasomes in the Lung [J]. FRONTIERS IN IMMUNOLOGY, 2020, 11.
[16] RAMIREZ M L G, POREBA M, SNIPAS S J, et al. Extensive peptide and natural protein substrate screens reveal that mouse caspase-11 has much narrower substrate specificity than caspase-1 [J]. JOURNAL OF BIOLOGICAL CHEMISTRY, 2018, 293(18): 7058-67.
[17] ABU KHWEEK A, AMER A O. Pyroptotic and non-pyroptotic effector functions of caspase-11 [J]. IMMUNOLOGICAL REVIEWS, 2020, 297(1): 39-52.
[18] ZHANG X, XU A, LV J, et al. Development of small molecule inhibitors targeting NLRP3 inflammasome pathway for inflammatory diseases [J]. EUROPEAN JOURNAL OF MEDICINAL CHEMISTRY, 2020, 185.
[19] VONG C T, TSENG H H L, YAO P, et al. Specific NLRP3 inflammasome inhibitors: promising therapeutic agents for inflammatory diseases [J]. DRUG DISCOVERY TODAY, 2021, 26(6): 1394-408.
[20] SEBASTIAN-VALVERDE M, WU H, AL RAHIM M, et al. Discovery and characterization of small-molecule inhibitors of NLRP3 and NLRC4 inflammasomes [J]. JOURNAL OF BIOLOGICAL CHEMISTRY, 2021, 296.
[21] HU J J, LIU X, XIA S, et al. FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation [J]. NATURE IMMUNOLOGY, 2020, 21(7): 736-+.
[22] PANDEYA A, LI L, LI Z, WEI Y. Gasdermin D (GSDMD) as a new target for the treatment of infection [J]. MEDCHEMCOMM, 2019, 10(5): 660-7.

