In indirect enzyme-linked immunosorbent assay (ELISA), the conformational state (native or denatured) of coated antigens serves as more than a simple experimental parameter; it acts as a fundamental systematic determinant that governs the entire assay process, from epitope recognition to final detection sensitivity. Native antigens primarily display conformational epitopes, while antigen denaturation exposes linear epitopes. These two types of epitopes exhibit distinct antibody binding affinities, which substantially alter the specificity of antibody antigen interactions. Meanwhile, denaturation induced changes in protein hydrophobicity and surface charge modulate the solid phase adsorption efficiency and molecular orientation of coated antigens, further restricting the accessibility of antigen epitopes to antibodies. In biomarker detection, denatured antigens can enhance the detection efficiency of certain low affinity antibodies by unmasking repetitive linear epitopes. However, the disruption of key conformational epitopes during denaturation may lead to the failure of capturing high affinity pathogenic antibodies, resulting in an inherent trade-off between assay sensitivity and specificity. This issue is particularly pivotal for autoantibody detection, as the binding of most disease associated autoantibodies depends on conformational epitope recognition. Accordingly, maintaining the native folding state of antigens is indispensable for accurately reflecting the in vivo immune status of patients. Furthermore, the exposure of cryptic epitopes triggered by antigen denaturation exerts dual effects on ELISA performance: it enables the capture of early stage and low abundance antibodies, yet it may also cause non-specific binding interference that compromises assay accuracy. Collectively, the conformational status of coated antigens profoundly affects all key steps of ELISA, including antigen coating, epitope recognition, antibody binding, and clinical result interpretation. A comprehensive understanding of the mechanistic effects of antigen conformational states provides a solid theoretical basis for the optimization of ELISA protocols and the accurate interpretation of inter sample variability.
This article reviews the differential recognition of linear versus conformational epitopes in indirect ELISA, compares the impact of protein denaturation on antigen coating efficiency, explains the shift in antibody specificity between native and denatured antigens, evaluates the discrepancies in biomarker detection sensitivity when using native versus denatured antigens, examines the role of protein conformation in ELISA based autoantibody detection, and addresses the exposure of cryptic epitopes by denatured proteins in immunoassays.
Table of Contents
1. Linear vs conformational epitope recognition in Indirect ELISA
2. Effect of protein denaturation on antigen coating efficiency
3. Antibody specificity shift between native and denatured antigens
4. Biomarker detection sensitivity with native vs denatured antigens
5. Role of protein conformation in autoantibody detection by ELISA
6. Denatured protein exposure of cryptic epitopes in immunoassays
01 Linear vs conformational epitope recognition in Indirect ELISA
The suitability of denatured antigens as coating substrates in indirect ELISA is fundamentally determined by the epitope type recognized by the corresponding antibodies, namely linear or conformational epitopes. A linear epitope (also referred to as a continuous epitope) consists of contiguous amino acid residues within an antigen’s primary sequence. Its recognition is independent of protein three-dimensional structure; therefore, antibodies targeting linear epitopes can retain binding capacity even after antigen denaturation, such as treatment with SDS-PAGE or alkaline reduction[1,2], while a conformational epitope (or discontinuous epitope) is assembled from amino acid residues that are far apart along the polypeptide backbone but spatially aggregated through protein folding, and antibody binding to such epitopes strictly depends on the maintenance of the antigen’s native tertiary structure[3], and during direct immobilization of antigens onto solid phase carriers for indirect ELISA, adsorption-induced conformational changes may mask conformational epitopes and abolish the binding of conformation specific antibodies, whereas antibodies that target linear epitopes are insensitive to such structural perturbations[4,5]; given this distinction, a standard experimental approach to characterize antibody epitope preference is to compare antibody ELISA reactivity against native (non-denatured) and denatured antigen preparations, whereby antibodies that exhibit exclusive reactivity to native antigens recognize conformational epitopes and antibodies binding equally to both antigen formats target linear epitopes[1,6], and for antibodies that solely recognize conformational epitopes, antigens with fully preserved native structures must be immobilized via alternative strategies such as capture antibody mediated indirect coating rather than direct passive adsorption, while linear epitope-directed antibodies tolerate denatured antigen substrates, a feature that forms the core principle of Western blotting, wherein target proteins are detected after separation via denaturing SDS-PAGE[2,7]; when constructing multi epitope recombinant antigens for broad spectrum detection of serum antibodies against various pathogens, tandem linear B cell epitopes are widely incorporated into antigen constructs, and this structural design ensures robust antibody recognition of diagnostic antigens despite partial unfolding occurring during antigen purification or plate coating procedures[8], collectively indicating that the practicality of using denatured coating antigens in indirect ELISA is governed by the epitope specificity of the test antibody: denatured antigens are applicable for detecting linear epitope targeted antibodies, whereas assays for conformational epitope specific antibodies necessitate antigens to maintain their native folded conformation.
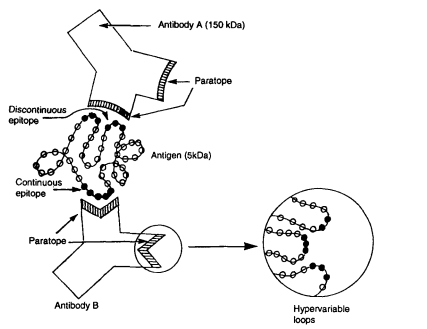
Fig. 1 Interaction of two antibodies with a continuous and a discontinuous epitope of an antigen[3].
02 Effect of protein denaturation on antigen coating efficiency
Protein denaturation exerts multifaceted and profound effects on coating efficiency and overall performance stability in ELISA. Despite being the most straightforward immobilization technique, direct passive adsorption frequently triggers adsorption mediated structural rearrangements in most coated antigens or antibodies, which compromise antigen binding capacity and disrupt intact epitope architecture[9,10]. Multiple investigations have corroborated this phenomenon; for example, antibodies immobilized on polystyrene microplate surfaces may lose over 90% of their functionally active binding sites following passive adsorption. In comparison, indirect immobilization strategies including streptavidin biotin coupling tether capture biomolecules to a pre coated intermediate layer, physically separating target proteins from hydrophobic solid phase substrates and substantially preserving their native folding and biological functionality[9,10]. Importantly, mild heat mediated partial denaturation does not universally impair coating performance. Relevant research has demonstrated that moderately heat denatured bovine serum albumin (BSA) assembles a more compact adsorbed film on silica matrices, exhibiting a 10 fold enhancement in blocking efficacy alongside accelerated adsorption kinetics. This favorable outcome is likely attributed to the unmasking of abundant hydrophobic domains that strengthen hydrophobic interactions between proteins and solid supports[11]. Analogously, thermal pre treatment reshapes the secondary and tertiary conformations of proteins such as β conglycinin during antigen coating by exposing additional hydrophobic moieties, thereby facilitating tighter attachment to the solid phase[12]. Nevertheless, extreme thermal denaturation irreversibly disrupts conformational epitopes, drastically diminishing the binding affinity of target specific immunoglobulins, particularly IgG molecules[11]. Accordingly, rational coating strategy selection necessitates a delicate balance between maximal adsorption yield and sustained epitope integrity: indirect immobilization or finely tuned partial denaturation is recommended for structurally labile antigens to elevate assay sensitivity and operational stability, while adequate blocking procedures remain indispensable for saturating unoccupied hydrophobic binding sites and mitigating non specific background signals[9,10]. Collectively, optimization of ELISA coating parameters requires comprehensive evaluation of intrinsic biochemical characteristics of antigens and antibodies, physicochemical properties of solid phase carriers, as well as the trade offs associated with protein structural unfolding. Data driven comparative assessment of diverse immobilization approaches and blocking regimens is therefore required to attain optimal analytical performance of immunoassays.
03 Antibody specificity shift between native and denatured antigens
The specificity of antibody binding varies substantially between native and denatured antigen conformations, representing a fundamental phenomenon that governs the interpretation of immunoassay outcomes and the rational selection of antibody reagents. Since conformational epitopes rely on the intact three-dimensional architecture of native proteins, they are readily disrupted during antigen denaturation, while linear epitopes are concomitantly exposed or newly generated[13,14]. This structural divergence leads to distinct antibody reactivity patterns: antibodies elicited against native proteins typically fail to bind their denatured counterparts, and antibodies induced by denatured antigens often exhibit negligible or no reactivity toward natively folded antigens[15,16]. A typical example is ovalbumin specific monoclonal antibodies, wherein antibodies derived from native ovalbumin predominantly target discontinuous conformational epitopes and display robust binding to native antigens but substantially diminished or abolished affinity for denatured ovalbumin, whereas antibodies raised against denatured ovalbumin recognize continuous linear epitopes and maintain comparable reactivity toward both native and denatured antigen forms. Notably, monoclonal antibodies screened against antigens directly immobilized on plastic surfaces tend to preferentially recognize partially or fully unfolded antigen variants rather than native conformations, primarily because passive adsorption during plate coating inherently induces protein structural perturbations[15,17]. Such conformational dependent specificity shifts are not merely theoretical observations but carry critical practical significance for immunoassay design, as antibodies with excellent performance in specific assay formats (e.g., standard ELISA with passively coated antigens) may be incompatible with applications requiring the recognition of soluble native proteins under physiological or in vivo conditions[16,18]. Therefore, rigorous characterization of antibody specificity toward both native and denatured antigens, via approaches including solution phase competition assays and capture ELISA using biotinylated antigens, is indispensable to validate the capacity of selected antibodies to faithfully reflect the intrinsic biological state of target proteins[16,17]. Comprehensive awareness of this specificity shift mechanism effectively prevents misleading experimental results and facilitates the targeted selection of qualified antibodies for specific application scenarios, whether for detecting native proteins in physiological specimens or identifying denatured epitopes in immunoblotting analyses.
04 Biomarker detection sensitivity with native vs denatured antigens
The selection of native versus denatured antigen formats exerts a profound impact on biomarker detection sensitivity within immunoassays, yet neither format holds universal superiority; instead, the optimal antigen configuration is dictated by the epitope specificity of the detection antibody and the adopted assay platform. Direct passive immobilization of native antigens on polystyrene microplate surfaces commonly triggers adsorption driven structural rearrangements that disrupt discontinuous conformational epitopes, resulting in drastically weakened antibody antigen interactions and compromised detection sensitivity[19,20]. As illustrated by Hnasko et al. (2011), numerous antibodies exhibiting robust performance in Western blotting an assay system relying on fully denatured antigens lose target recognition capability in conventional indirect ELISA employing natively coated antigens; nevertheless, brief chemical denaturation treatment with guanidine hydrochloride boosted detection sensitivity for multiple brain biomarkers (including glutamine synthetase, OMG, and BAI-1) by as much as 10⁷ fold, a phenomenon attributable to the unmasking of linear epitopes that remain concealed within the folded native protein structure. On the contrary, antibodies generated against intact native proteins predominantly target conformation dependent discontinuous epitopes, which become irreversibly disrupted following denaturation; these antibodies display potent binding affinity toward native antigens yet show drastically attenuated or even undetectable reactivity against denatured protein substrates[16,17]. For clinical biomarker testing, coating plates with denatured antigens accordingly improves analytical sensitivity for linear epitope specific antibodies, whereas native antigen immobilization is mandatory for capturing antibodies that recognize conformational epitopes a consideration of great clinical relevance for diagnostic platforms in which biological specimens contain biomarkers in their physiological native state[16,17]. Additionally, matrix constituents and coexisting interfering proteins serve as additional confounding variables shaping overall detection sensitivity: Kenny and Dunsmoor (1983) verified that abundant competing proteins within mixed antigen samples hinder efficient immobilization of target analytes, particularly when the target protein accounts for less than 1% of the total protein pool, ultimately diminishing assay sensitivity. Furthermore, the analytical performance obtained with denatured antigen coatings can be further refined through rational optimization of experimental parameters, including the choice of denaturing agent (guanidine hydrochloride confers superior epitope exposure compared to urea or SDS), modulation of denaturant concentration and incubation duration, and utilization of high binding capacity polystyrene microplates. Collectively, biomarker detection sensitivity is not an inherent characteristic of a given antigen but arises from the synergistic interplay of three core factors: antigen conformational status (native or denatured), antibody epitope specificity, and overall immunoassay design. For this reason, parallel empirical evaluation of both native and denatured antigen preparations is strongly advised to establish the most sensitive testing scheme for each unique antibody target biomarker pair[16,17,20].
05 Role of protein conformation in autoantibody detection by ELISA
For ELISA based autoantibody testing, the conformational status of capture antigens constitutes a decisive experimental variable, given that the vast majority of clinically meaningful autoantibodies target discontinuous conformational epitopes exclusively displayed by natively folded proteins, structural motifs that become irreversibly disrupted after protein denaturation[21,22]. Autoantibodies against Ro/SSA and Jo-1 serve as representative examples of conformation dependent immunoreactivity; reliable ELISA quantification of these autoantibodies strictly relies on intact native antigens, whereas denatured antigen substrates frequently produce false negative diagnostic readouts. Likewise, anti double stranded DNA (dsDNA) autoantibodies, a serological hallmark with high diagnostic specificity for systemic lupus erythematosus (SLE), recognize unique helical structural signatures exclusive to native double stranded DNA. Substitution with single stranded DNA or misfolded surface adsorbed dsDNA triggers non specific cross reactivity with antibodies lacking clinical disease correlation[21,22]. A major technical limitation of traditional ELISA lies in passive protein adsorption onto polystyrene microplates, a procedure that frequently triggers adsorption mediated protein unfolding and abolishes critical conformational epitopes, ultimately impairing both analytical sensitivity and diagnostic specificity. Capture ELISA has emerged as an effective workaround to mitigate this artifact: this platform pre coats wells with monoclonal capture antibodies to spatially orient target antigens and sustain their native tertiary architecture, thereby maintaining unobstructed access of autoantibodies to intact conformational epitopes. Additionally, antigen conformation exerts secondary effects on two key assay parameters: the selection of detection conjugates (IgG specific vs. Pan immunoglobulin reagents) and the establishment of diagnostic cut off thresholds, owing to divergent binding affinities exhibited by distinct immunoglobulin subclasses toward native and denatured antigen conformations. Although denatured antigens and synthetic peptide substrates are routinely implemented in Western blotting and peptide immunoassays to interrogate linear epitopes, such platforms are inappropriate for quantifying most conformation dependent autoantibodies, and their measured signals often fail to align with in vivo clinical disease activity. Collectively, robust and clinically translatable autoantibody serodiagnosis mandates the utilization of structurally preserved native antigens. Comprehensive optimization of ELISA workflows, covering antigen purification protocols, biomolecule immobilization schemes, and buffered incubation environments, is therefore indispensable to safeguard the integrity of disease relevant conformational epitopes.
06 Denatured protein exposure of cryptic epitopes in immunoassays
Protein denaturation enables the exposure of cryptic epitopes segments concealed within the native three dimensional protein structure and inherently inaccessible to antibodies thereby fundamentally reshaping the outcomes of immunoassays[15,23]. Friguet et al. (1984) validated this mechanism using monoclonal antibodies generated against the native β2 subunit of Escherichia coli tryptophan synthase: certain antibodies exhibited extremely slow binding kinetics toward soluble native proteins but rapidly recognized denatured antigens in ELISA competition assays, confirming that these epitopes remain structurally masked in the native conformation and become surface exposed only upon protein unfolding. This represents a prevalent rather than sporadic phenomenon; subsequent studies have verified that numerous monoclonal antibodies screened against surface adsorbed antigens, which frequently undergo partial denaturation during immobilization, preferentially target cryptic linear epitopes instead of native conformational epitopes. The biological functions of such cryptic epitopes have attracted growing research attention. Mortimer and Minchin (2016) summarized a broad spectrum of extracellular matrix proteins, including collagen, fibronectin, laminin, and fibrinogen, that contain buried “matricryptic sites”; these latent sequences are unmasked following conformational alterations induced by mechanical stretching, thermal denaturation, or nanoparticle binding, and subsequently mediate diverse physiological activities such as integrin interaction, cell adhesion, and angiogenesis. In immunoassay workflows, deliberate or inadvertent protein denaturation during antigen coating and sample preparation facilitates the detection of antibodies targeting newly exposed linear cryptic epitopes, while completely abrogating the binding reactivity of conformational epitope specific antibodies. For instance, Hnasko et al. (2011) demonstrated that chemical denaturation via guanidine hydrochloride treatment enhanced the indirect ELISA detection sensitivity of multiple brain biomarkers by up to 10⁷ fold, an improvement attributed to the unmasking of linear cryptic epitopes hidden in native protein structures. Consistent with these findings, It is reported distinct secondary structural characteristics in differently prepared aflatoxin B1 protein conjugate coating antigens, as verified by circular dichroism analysis, and such structural variations were strongly correlated with differences in ELISA sensitivity and antibody titer, further demonstrating that epitope accessibility is strictly governed by the conformational status of immobilized antigens[24]. In contrast, for autoimmune diagnostic applications, the utilization of denatured antigens frequently causes false negative results, as disease relevant autoantibodies targeting conformation dependent epitopes fail to recognize structurally disrupted denatured antigens. Collectively, protein denaturation does not merely impair antigenicity but induces a comprehensive shift in the antigen epitope repertoire from conformational to linear epitope dominance, which either improves or compromises immunoassay performance depending on the epitope specificity of target antibodies. Accordingly, the design and result interpretation of immunoassays must fully account for antigen conformational status (native, partially denatured, or fully unfolded), which defines the profile of accessible epitopes. The exposure of cryptic epitopes can serve as a powerful strategy to enhance the detection of linear epitope specific antibodies, yet it may also introduce analytical artifacts when the antigen conformational state is mismatched with the assay’s intended clinical or research application[25].
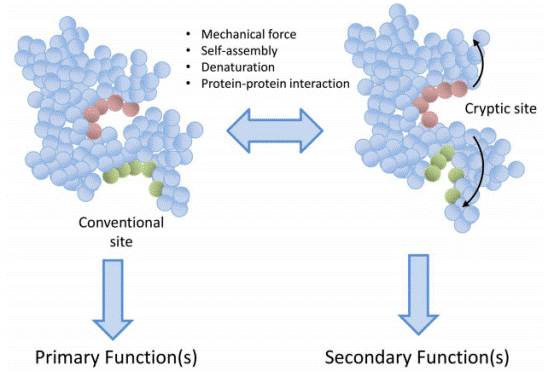
Fig. 2 Cryptic epitopes. Representation of how buried sequences can be uncovered following unfolding induced by various factors such as mechanical forces or protein protein interactions. Primary function of the protein is attributed to sites accessible when the protein is in its normal folded state. Secondary functions, which may be entirely different to the primary function, are only revealed following unfolding. Importantly, this process is reversible[23].
In summary, the conformational status of coated antigens (native versus denatured) represents a critical systematic determinant rather than a negligible experimental variable in indirect ELISA, profoundly modulating the entire assay workflow from epitope accessibility to the final interpretation of diagnostic results. Natively folded antigens retain intact conformational epitopes, which are indispensable for capturing high affinity and clinically significant antibodies, especially in autoantibody detection; nevertheless, such antigens are susceptible to adsorption induced structural disruption during passive immobilization, which inevitably compromises assay sensitivity. By contrast, denatured antigens unmask linear and frequently cryptic epitopes, enabling substantial improvements in the detection of low abundance antibodies and antibodies targeting linear epitopes, though this structural alteration may lead to the failure of identifying conformation dependent antibodies and introduce undesirable non specific interference. Furthermore, denaturation remodels protein hydrophobicity and surface binding characteristics, thereby altering antigen coating efficiency and molecular orientation, while antibody binding specificity can differ drastically between native and denatured antigen formats, highlighting the necessity of rigorous antibody reagent characterization. Notably, neither native nor denatured antigen configuration is universally superior, as the optimal antigen format is contingent on the epitope specificity of target antibodies, the adopted assay platform, and the intended research or clinical application. Parallel empirical evaluation of both native and denatured antigen preparations, together with the optimization of antigen immobilization strategies and blocking conditions, is therefore essential to achieve an optimal balance between assay sensitivity and specificity. Overall, mechanistic insight into how antigen conformation regulates epitope presentation, antibody antigen recognition, and solid phase protein interactions lays a solid theoretical basis for the rational design of ELISA protocols and the reliable interpretation of sample to sample variability.
Elabscience® Quick Overview of Popular Products:
Table 1. Research Tools for Biomarker Detection
|
Cat. No. |
Product Name |
|
E-TO-E017 |
AFB1(Aflatoxin B1) ELISA Kit |
|
E-EL-R2494 |
Rat GS(Glutamine synthetase) ELISA Kit |
|
E-AB-16668 |
OMG Polyclonal Antibody |
|
E-EL-H0169 |
Human IgG(Immunoglobulin G) ELISA Kit |
References:
[1] Christensen, N. D., Dillner, J., Eklund, C., Carter, J. J., Wipf, G. C., Reed, C. A., Cladel, N. M., & Galloway, D. A. (1996). Surface conformational and linear epitopes on HPV-16 and HPV-18 L1 virus-like particles as defined by monoclonal antibodies. Virology,223(1), 174–184. https://doi.org/10.1006/viro.1996.0466
[2] Forsström, B., Axnäs, B. B., Rockberg, J., Danielsson, H., Bohlin, A., & Uhlen, M. (2015). Dissecting antibodies with regards to linear and conformational epitopes.PloS one,10(3), e0121673. https://doi.org/10.1371/journal.pone.0121673
[3] van Regenmortel M. H. (2000). The recognition of proteins and peptides by antibodies.Journal of immunoassay,21(2-3), 85–108. https://doi.org/10.1080/01971520009349530
[4] Dekker, E. L., Porta, C., & Van Regenmortel, M. H. (1989). Limitations of different ELISA procedures for localizing epitopes in viral coat protein subunits.Archives of virology, 105(3-4), 269–286. https://doi.org/10.1007/BF01311363
[5] Kenett D. (1988). A simple ELISA for the classification of monoclonal antibodies according to their recognition of native epitopes. Journal of immunological methods, 106(2), 203–209. https://doi.org/10.1016/0022-1759(88)90198-6
[6] Manaresi, E., Zuffi, E., Gallinella, G., Gentilomi, G., Zerbini, M., & Musiani, M. (2001). Differential IgM response to conformational and linear epitopes of parvovirus B19 VP1 and VP2 structural proteins. Journal of medical virology, 64(1), 67–73. https://doi.org/10.1002/jmv.1019
[7] Riddell, M. A., Li, F., & Anderson, D. A. (2000). Identification of immunodominant and conformational epitopes in the capsid protein of hepatitis E virus by using monoclonal antibodies. Journal of virology, 74(17), 8011–8017.https://doi.org/10.1128/jvi.74.17.8011-8017.2000
[8] Liu, L., Chen, Q., Yu, L., Wu, W., Mou, S., Guo, H., Tan, C., Liu, Q., Wang, D., Chen, H., & Wang, X. (2026). Development of an indirect ELISA based on a multi-epitope recombinant protein for antibody detection against African swine fever virus. Microbiology spectrum, e0352525. Advance online publication. https://doi.org/10.1128/spectrum.03525-25
[9] Butler J. E. (2000). Solid supports in enzyme-linked immunosorbent assay and other solid-phase immunoassays. Methods (San Diego, Calif.), 22(1), 4–23. https://doi.org/10.1006/meth.2000.1031
[10] Näreoja, T., Määttänen, A., Peltonen, J., Hänninen, P. E., & Härmä, H. (2009). Impact of surface defects and denaturation of capture surface proteins on nonspecific binding in immunoassays using antibody-coated polystyrene nanoparticle labels. Journal of immunological methods, 347(1-2), 24–30. https://doi.org/10.1016/j.jim.2009.05.010
[11] Park, J. H., Jackman, J. A., Ferhan, A. R., Ma, G. J., Yoon, B. K., & Cho, N. J. (2018). Temperature-Induced Denaturation of BSA Protein Molecules for Improved Surface Passivation Coatings. ACS applied materials & interfaces, 10(38), 32047–32057. https://doi.org/10.1021/acsami.8b13749
[12] Li, T., Bu, G., & Xi, G. (2021). Effects of heat treatment on the antigenicity, antigen epitopes, and structural properties of β-conglycinin. Food chemistry, 346, 128962. https://doi.org/10.1016/j.foodchem.2020.128962
[13] Abbott, W. M., Damschroder, M. M., & Lowe, D. C. (2014). Current approaches to fine mapping of antigen-antibody interactions. Immunology, 142(4), 526–535. https://doi.org/10.1111/imm.12284
[14] Braden, B. C., & Poljak, R. J. (1995). Structural features of the reactions between antibodies and protein antigens. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 9(1), 9–16. https://doi.org/10.1096/fasebj.9.1.7821765
[15] Friguet, B., Djavadi-Ohaniance, L., & Goldberg, M. E. (1984). Some monoclonal antibodies raised with a native protein bind preferentially to the denatured antigen. Molecular immunology, 21(7), 673–677. https://doi.org/10.1016/0161-5890(84)90053-1
[16] Holm, B. E., Bergmann, A. C., Hansen, P. R., Koch, C., Houen, G., & Trier, N. H. (2015). Antibodies with specificity for native and denatured forms of ovalbumin differ in reactivity between enzyme-linked immunosorbent assays. APMIS : acta pathologica, microbiologica, et immunologica Scandinavica, 123(2), 136–145. https://doi.org/10.1111/apm.12329
[17] Koch, C., Jensen, S. S., Oster, A., & Houen, G. (1996). A comparison of the immunogenicity of the native and denatured forms of a protein. APMIS : acta pathologica, microbiologica, et immunologica Scandinavica, 104(2), 115–125. https://doi.org/10.1111/j.1699-0463.1996.tb00696.x
[18] Gao, J., Sidhu, S. S., & Wells, J. A. (2009). Two-state selection of conformation-specific antibodies. Proceedings of the National Academy of Sciences of the United States of America, 106(9), 3071–3076. https://doi.org/10.1073/pnas.0812952106
[19] Kenny, G. E., & Dunsmoor, C. L. (1983). Principles, problems, and strategies in the use of antigenic mixtures for the enzyme-linked immunosorbent assay. Journal of clinical microbiology, 17(4), 655–665. https://doi.org/10.1128/jcm.17.4.655-665.1983
[20] Hnasko, R., Lin, A., McGarvey, J. A., & Stanker, L. H. (2011). A rapid method to improve protein detection by indirect ELISA. Biochemical and biophysical research communications, 410(4), 726–731. https://doi.org/10.1016/j.bbrc.2011.06.005
[21] Tozzoli, R., Bizzaro, N., Tonutti, E., Villalta, D., Bassetti, D., Manoni, F., Piazza, A., Pradella, M., Rizzotti, P., & Italian Society of Laboratory Medicine Study Group on the Diagnosis of Autoimmune Diseases (2002). Guidelines for the laboratory use of autoantibody tests in the diagnosis and monitoring of autoimmune rheumatic diseases. American journal of clinical pathology, 117(2), 316–324. https://doi.org/10.1309/Y5VF-C3DM-L8XV-U053
[22] Meroni, P. L., Biggioggero, M., Pierangeli, S. S., Sheldon, J., Zegers, I., & Borghi, M. O. (2014). Standardization of autoantibody testing: a paradigm for serology in rheumatic diseases. Nature reviews. Rheumatology, 10(1), 35–43. https://doi.org/10.1038/nrrheum.2013.180
[23] Mortimer, G. M., & Minchin, R. F. (2016). Cryptic epitopes and functional diversity in extracellular proteins. The international journal of biochemistry & cell biology, 81(Pt A), 112–120. https://doi.org/10.1016/j.biocel.2016.10.020
[24] Ye, Y., Liu, A., Wang, X., & Chen, F. (2016). Spectra analysis of coating antigen: a possible explanation for difference in anti-AFB1 polyclonal antibody sensitivity. Journal of Molecular Structure, 1121, 74-79
[25] Kelker, H. C., Itri, V. R., & Valentine, F. T. (2010). A strategy for eliciting antibodies against cryptic, conserved, conformationally dependent epitopes of HIV envelope glycoprotein. PloS one, 5(1), e8555. https://doi.org/10.1371/journal.pone.0008555

