The imbalance between regulatory T cells (Tregs) and T helper 17 cells (Th17) has been increasingly recognized as a key immunoregulatory mechanism in gestational diabetes mellitus (GDM).
This review systematically summarizes the interactions between placental macrophages and Treg/Th17 cells involved in inflammation related to gestational diabetes, relevant mouse models applied to explore Treg/Th17-mediated placental inflammatory processes, as well as co-culture models (Treg/Th17 co-culture model) of trophoblasts and Treg/Th17 cells for exploring maternal-fetal immune imbalance under such diabetic conditions. It also elaborates on the in vitro regulatory mechanism whereby hyperglycemia drives naive CD4+ T cells to differentiate into Th17 cells, meanwhile introducing flow cytometry approaches used to detect the Treg/Th17 ratio in peripheral blood and decidual tissues from patients with gestational diabetes, alongside ELISA and multi-index cytokine detection methods adopted to monitor alterations in typical inflammatory cytokines linked to Treg/Th17 imbalance.
Table of Contents
1. Placental macrophage interactions with Treg/Th17 cells in gestational diabetes-associated inflammation
2. Mouse models of gestational diabetes for investigating Treg/Th17-mediated placental inflammation
3. Co-culture models of trophoblasts and Treg/Th17 cells for studying maternal-fetal immune imbalance in gestational diabetes
4. Hyperglycemia-induced differentiation of naïve CD4+ T cells toward the Th17 phenotype in in vitro gestational diabetes models
5. Flow cytometric strategies for quantifying the Treg/Th17 ratio in peripheral blood and decidual tissues of gestational diabetes patients
6. ELISA and multiplex cytokine panels for monitoring IL-10, IL-17A, and TGF-β changes associated with Treg/Th17 imbalance in gestational diabetes
01 Placental macrophage interactions with Treg/Th17 cells in gestational diabetes-associated inflammation
Gestational diabetes mellitus (GDM) is a common metabolic complication of pregnancy defined by glucose intolerance. GDM elicits systemic and local inflammation that profoundly affects the maternal-fetal interface, especially the placenta[1]. This inflammatory milieu acts as a core driver in GDM pathogenesis and is tightly associated with aberrations in immune cell populations, notably placental macrophages and the balance between regulatory T (Treg) and T helper 17 (Th17) cells[2].
Placental macrophages, consisting of maternal decidual macrophages and fetal Hofbauer cells (HBCs), are highly plastic immune cells essential for immune tolerance, host defense, and placental development. In healthy pregnancies, decidual macrophages predominantly display a non-inflammatory, pro-repair M2-like phenotype that supports the establishment and maintenance of maternal-fetal tolerance. In GDM, however, these cells frequently undergo a phenotypic shift toward a pro-inflammatory M1 state. Such altered polarization enhances the secretion of pro-inflammatory cytokines, including tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and interleukin-1β (IL-1β). Consistently, studies have reported elevated placental levels of TNF-α and IL-6 in women with GDM. This pro-inflammatory cascade is further reinforced by upregulated miR-185-5p, which promotes placental inflammation in GDM via targeting CDC42. Collectively, these changes establish the chronic low-grade inflammation characteristic of GDM[3,4].
The crosstalk between placental macrophages and T-cell subsets, particularly Tregs and Th17 cells, is indispensable for preserving immune homeostasis at the maternal-fetal interface. Tregs are central regulators of maternal-fetal immune tolerance. They suppress excessive immune reactivity through the secretion of anti-inflammatory cytokines such as interleukin-10 (IL-10) and transforming growth factor-beta (TGF-β), as well as by IL-2 consumption. Induced regulatory T cells (iTregs), derived from naive CD4+ T cells, are highly enriched in the decidua and play a pivotal role in preventing fetal rejection by establishing a local immune-privileged niche. In contrast, Th17 cells represent a pro-inflammatory T-cell subset that secretes IL-17A and exacerbates inflammatory responses[5].
GDM is associated with marked dysregulation of the Th17/Treg balance, typically manifested by reduced Treg frequency and function and an increased Th17/Treg ratio. This imbalance disrupts the fine immune tolerance required for normal pregnancy. Compared with healthy pregnant women, women with GDM exhibit lower IL-10 and higher IL-17A levels in peripheral blood mononuclear cells (PBMCs). This skewed phenotype is driven primarily by pro-inflammatory cytokines secreted by activated placental macrophages. For example, IL-6 promotes Th17 differentiation, whereas diminished IL-10 and TGF-β impair Treg development[6,7].
Hyperglycemia and elevated free fatty acids, key metabolic features of GDM, activate the TLR4/NF-κB signaling pathway in placental macrophages, further amplifying the release of IL-6 and TNF-α. In turn, these cytokines stimulate the IL-6–STAT3–RORγt axis to drive Th17 expansion, while suppressing the IL-2–STAT5–FOXP3 pathway critical for Treg differentiation and function. The consequent decline in Treg activity and expansion of Th17 cells contribute to impaired immune tolerance, heightened inflammation, and insulin resistance in GDM[8,9].
Notably, obesity, a frequent comorbidity of GDM, can independently trigger subclinical inflammation. Immunohistochemical analyses of placental macrophages in obese women with GDM reveal phenotypic and functional imbalance. Inflammation driven by GDM extends beyond the maternal-fetal interface and may compromise other immune defenses, including specific humoral and cellular responses against pathogens such as Toxoplasma gondii[10,11].
The inflammatory state in GDM also contributes to adverse pregnancy outcomes. Placental inflammation acts as a key mediator of maternal and neonatal complications. Emerging evidence indicates that GDM increases susceptibility to Group B Streptococcus (GBS) infection by impairing maternal immunity and disrupting vaginal microbiota, thereby increasing the risk of GBS transmission and poor neonatal outcomes[12]. Altered cytokine profiles in GDM placentas, particularly elevated IL-1β and activation of Toll-like receptor signaling, underscore the complex inflammatory network involved. Moreover, metabolic reprogramming in the GDM placenta, including disturbed lipid metabolism and impaired amino acid transport, is closely linked to inflammatory cytokine activity and mitochondrial stress[13].
Targeting inflammatory pathways and restoring the Treg/Th17 balance represents a promising therapeutic strategy for GDM. For example, curcumin modulates inflammatory responses in PBMCs from women with GDM by regulating the Treg/Th17 axis and associated cytokines (IL-17A, IL-6, and IL-10). Chitosan-low-molecular-weight heparin sodium nanoparticles have been shown to normalize the Treg/Th17 balance and alleviate inflammation in preeclampsia, suggesting a comparable therapeutic potential in GDM. TGF-β, a pleiotropic cytokine, critically modulates maternal-fetal crosstalk by shaping the placental immunomodulatory secretome, and its dysregulation is implicated in various pregnancy complications[14].
In conclusion, GDM is characterized by pronounced placental inflammation driven by macrophage polarization and an imbalanced Treg/Th17 axis. The pro-inflammatory milieu triggered by hyperglycemia and fatty acids promotes M1 macrophage activation and Th17 dominance, thereby impairing maternal-fetal immune tolerance and contributing to adverse pregnancy outcomes. A deeper understanding of these immune interactions is critical for developing targeted interventions to alleviate the detrimental effects of GDM on maternal and fetal health.
_.png)
Fig. 1 Detection of Th17 cells in human peripheral blood mononuclear cells (PBMCs). Peripheral blood mononuclear cells (PBMCs) were stimulated with Cell Stimulation MIX (Cat. No. E-CK-A019) and Protein Transport Inhibitor MIX (Cat. No. E-CK-A013) for 5 hours. After harvesting, fixation and permeabilization, cells were stained with PerCP/Cyanine5.5 anti-human CD3, Elab Fluor® 488 anti-human CD4 and PE anti-human IL-17A. The proportion of Th17 cells and their ability to secrete IL-17A were analyzed. Th17 cells were identified as CD3+CD4+IL-17A+ cells. (The data are provided by Elabscience.)
Elabscience® Quick Overview of Popular Products:
Table 1. Multicolor Panel for Flow Cytometric Analysis of Human and Mouse Helper T cells (CD4+ T cells)
|
Product Name |
Clone |
Fluorochrome |
Cat. No. |
Species Reactivity |
|
CD3 |
OKT-3 |
PerCP/Cyanine5.5 |
E-AB-F1001J |
Human |
|
CD4 |
SK3 |
Elab Fluor® 488 |
E-AB-F1352L |
Human |
|
IL-17A |
BL168 |
PE |
E-AB-F1173D |
Human |
|
IFN-γ |
B27 |
APC |
E-AB-F1196E |
Human |
|
IL-4 |
MP4-25D2 |
PE |
E-AB-F1203D |
Human |
|
CD3 |
17A2 |
PE/Cyanine5 |
E-AB-F1013G |
Mouse |
|
CD4 |
GK1.5 |
FITC |
E-AB-F1097C |
Mouse |
|
IFN-γ |
XMG1.2 |
APC |
E-AB-F1101E |
Mouse |
|
IL-4 |
11B11 |
PE |
E-AB-F1204D |
Mouse |
|
IL-17A |
17F3 |
PE |
E-AB-F1272D |
Mouse |
02 Mouse models of gestational diabetes for investigating Treg/Th17-mediated placental inflammation
Mouse models are indispensable tools for investigating the intricate interplay between Treg/Th17 cells and placental inflammation in gestational diabetes mellitus (GDM), offering controlled environments to dissect pathophysiological mechanisms that are challenging to study in human pregnancies. The ideal GDM mouse model should accurately mimic the key clinical and pathophysiological features of human GDM, including hyperglycemia, insulin resistance, and the associated inflammatory responses at the maternal-fetal interface[15].
Various approaches are employed to establish GDM in mice, each with distinct advantages and limitations for studying Treg/Th17-mediated placental inflammation:
Diet-induced animal models: Diet-induced animal models, including high-fat diet (HFD) and high-fat high-sugar (HFHS) diet, are widely used to establish obesity-related insulin resistance and hyperglycemia that recapitulate key features of human gestational diabetes mellitus (GDM). These dietary models trigger systemic inflammation, disrupt Treg/Th17 cell balance, impair placental function and glucose metabolism, and ultimately generate a GDM-like phenotype[16]. In contrast, high fermentable dietary fiber (HFDF) intervention represents a potential strategy to alleviate GDM-associated placental inflammation by regulating the gut flora–short-chain fatty acid-placental inflammation axis[17].
Chemically-Induced Models: Streptozotocin (STZ) induces gestational diabetes mellitus (GDM) by selectively impairing pancreatic beta-cell function and causing mild hyperglycemia without inducing severe diabetes, thus maintaining fetal survival. Administration of STZ (e.g., 75 mg/kg for two consecutive days before pregnancy) in C57BL/6N mice effectively generates a stable GDM model that supports detailed phenotypic analysis of disease progression. This model has been widely used to investigate the contributions of inflammation and specific cytokines such as interleukin-1β (IL-1β) to GDM pathogenesis. Notably, inhibition of IL-1β has been reported to improve glycemic control in STZ-induced GDM mice, supporting the critical involvement of inflammatory signaling in GDM development[18].
Genetic Models: While specific genetic models like db/db or ob/ob mice can exhibit severe GDM phenotypes, their use in studying Treg/Th17-mediated placental inflammation can be complicated by embryonic lethality or severe fetal growth restriction. However, carefully designed conditional or heterozygous crosses allow for more controlled investigations. Genetic predisposition is also considered a risk factor for GDM. Dendrobine has been shown to attenuate GDM in a genetic mouse model by inhibiting Th17 cells, suggesting the relevance of these models for therapeutic studies[19].
Fecal Microbiota Transplantation (FMT) Models: Recent studies highlight the role of gut microbiota dysbiosis in GDM. FMT from GDM patients to healthy mice has been demonstrated to induce glucose intolerance and subclinical inflammation, mimicking GDM-like phenotypes. This innovative approach allows for the investigation of gut-placenta axis interactions in GDM-associated inflammation and immune cell dysregulation[20].
Mouse models have revealed that GDM skews the decidual Treg/Th17 balance towards Th17 dominance, promoting pro-inflammatory cytokine production, oxidative stress, and impaired spiral artery remodeling–mechanisms linked to placental dysfunction and adverse pregnancy outcomes. For example, the impaired balance of Tregs and Th17 cells is also linked to preeclampsia, where Yin Yang 1 (YY1) has been shown to impact preeclampsia by regulating Treg/Th17 cells and the PI3K/AKT pathway[21]. The critical role of the placenta as an immuno-endocrine organ that recognizes and responds to metabolic distress is underscored by these findings. Furthermore, specific alterations in inflammatory profiles have been observed in tissues affected by GDM.
These models have also enabled the evaluation of potential therapeutic interventions. Anakinra, an IL-1 receptor antagonist, has been shown to ameliorate insulin resistance and inflammation in a murine GDM model by downregulating Th17 responses. Another study demonstrated that dendrobine attenuates GDM in mice by inhibiting Th17 cells. These findings highlight the utility of mouse models for identifying and validating therapeutic targets for GDM and its associated inflammatory complications.
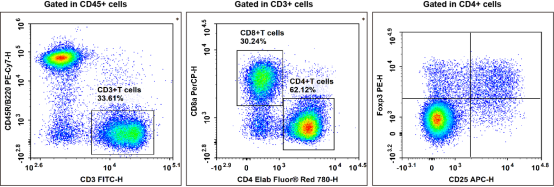
Fig. 2 Detection and analysis of Treg cells in C57BL/6 mouse splenocytes. C57BL/6 mouse splenocytes were surface stained with Elab Fluor® Violet 450 Anti-Mouse CD45, PE/Cyanine7 Anti-Mouse CD45R/B220, FITC Anti-Mouse CD3, Elab Fluor® Red 780 Anti-Mouse CD4, PerCP Anti-Mouse CD8a and APC Anti-Mouse CD25 and then treated with Foxp3/Transcription Factor Staining Kit. Cells were then stained with PE Anti-Mouse Foxp3, followed by analysis via flow cytometry. Regulatory T cells (Treg cells) exhibit the phenotype of CD45+CD45R/B220-CD3+CD4+CD25+Foxp3+. (The data are provided by Elabscience.)
Elabscience® Quick Overview of Popular Products:
Table 2. Reagents for Mouse Treg cell Research
|
Product Name |
Cat. No. |
|
Elab Fluor® Violet 450 Anti-Mouse CD45 Antibody[30-F11] |
E-AB-F1136Q |
|
PE/Cyanine7 Anti-Mouse CD45R/B220 Antibody[RA3.3A 1/6.1] |
E-AB-F1112H |
|
FITC Anti-Mouse CD3 Antibody[17A2] |
E-AB-F1013C |
|
Elab Fluor® Red 780 Anti-Mouse CD4 Antibody[RM4-5] |
E-AB-F1353S |
|
PerCP Anti-Mouse CD8a Antibody[53-6.7] |
E-AB-F1104F |
|
APC Anti-Mouse CD25 Antibody[PC-61.5.3] |
E-AB-F1102E |
|
PE Anti-Mouse Foxp3 Antibody[3G3] |
E-AB-F1238D |
|
10×ACK Lysis Buffer |
E-CK-A105 |
|
Ready-to-Use Foxp3/Transcription Factor Staining Kit |
E-CK-A108 |
03 Co-culture models of trophoblasts and Treg/Th17 cells for studying maternal-fetal immune imbalance in gestational diabetes
Co-culture models (Treg/Th17 co-culture model) serve as robust in vitro tools to explore the complex cellular crosstalk between trophoblasts and immune cells, especially regulatory T cells (Tregs) and T helper 17 (Th17) cells, in the setting of maternal-fetal immune imbalance induced by gestational diabetes mellitus (GDM). These models enable researchers to establish controllable experimental conditions and further clarify the molecular mechanisms underlying GDM-related placental inflammation and immune dysfunction, which are difficult to systematically investigate through in vivo research[22].
3.1 Rationale for Establishing Co-culture Models (Treg/Th17 co-culture model) in GDM Research
As an organ integrating immune and endocrine functions, the placenta is core to sensing and responding to metabolic stress such as that occurring under GDM conditions. GDM is featured by chronic low-grade inflammation at the maternal-fetal interface, accompanied by disordered cytokine profiles and abnormal immune cell functions. The dysregulation of the Treg/Th17 axis, a pivotal factor for maintaining maternal-fetal immune tolerance, constitutes a key manifestation of such immune imbalance. As the predominant epithelial cells in the placenta, trophoblasts lie at the core position of maternal-fetal interaction and actively communicate with immune cells residing in the decidua and placental tissues. Accordingly, co-culturing trophoblasts with Tregs and Th17 cells helps clarify how GDM-specific pathological microenvironments modulate such intercellular crosstalk and further drive the progression of placental inflammation[23,24].
3.2 Establishment of Co-culture Models (Treg/Th17 co-culture model)
(1) Selection and Treatment of Trophoblasts
Researchers can adopt human trophoblast cell lines including HTR-8/SV40, BeWo and JAR, or primary trophoblasts isolated from placental tissues for co-culture experiments. Primary trophoblasts are preferred owing to their superior similarity to in vivo physiological states, despite greater difficulties in isolation and culture procedures. When applying primary cells, researchers must strictly take the gestational age and GDM status of placental donors into account to accurately restore physiological or pathological characteristics. In addition, trophoblasts can be pretreated with high glucose, high free fatty acids or pro-inflammatory cytokines such as IL-1β, TNF-α and IL-6 to simulate the GDM microenvironment, which facilitates exploring the direct impacts of metabolic stressors on trophoblast biological functions and their subsequent interactions with immune cells[24,25].
(2) Isolation, Expansion and Identification of Tregs and Th17 Cells
Tregs and Th17 cells can be isolated from peripheral blood mononuclear cells (PBMCs) or cord blood samples collected from GDM patients and healthy pregnant women via magnetic bead sorting or flow cytometry according to specific surface markers, namely CD4, CD25 and CD127 for Tregs and CD4 for Th17 cells. Alternatively, induced Tregs and Th17 cells can be generated from naive CD4+ T cells through in vitro differentiation to obtain stable and standardized cell sources. After isolation and expansion, flow cytometry is required for rigorous phenotypic identification by detecting core transcription factors including FOXP3 in Tregs and RORγt in Th17 cells, as well as characteristic cytokines secreted by these cells, so as to verify cell identity and guarantee complete biological functions[26].
(3) Construction of Co-culture Systems
Two mainstream co-culture patterns are commonly applied in relevant studies. Direct co-culture allows intimate contact between trophoblasts and T cells, which is suitable for exploring direct intercellular communication and ligand-receptor interactions. By contrast, indirect co-culture systems represented by Transwell chambers separate two types of cells with porous membranes to block direct cell contact while permitting the exchange of soluble substances, thus being ideal for analyzing the regulatory roles of secreted cytokines and chemokines. The optimal proportion of trophoblasts to T cells needs to be determined through pre-experiments, and the commonly adopted ratio ranges from 1:1 to 1:10, which can well simulate in vivo physiological ratios and amplify observable cellular response phenotypes[27].
To comprehensively evaluate Treg/Th17-mediated placental inflammatory responses using GDM co-culture models, multiple core detection indicators should be analyzed.
(1) Dynamic Changes in Treg/Th17 Balance
Flow cytometry is used to quantify the proportion of CD4+FOXP3+ Tregs and CD4+RORγt+IL-17A+ Th17 cells after co-culture to reflect their numerical alterations, and an elevated Th17/Treg ratio is regarded as a typical sign of GDM-related maternal-fetal immune imbalance. Meanwhile, quantitative real-time PCR can detect the mRNA expression levels of key functional genes such as FOXP3, RORC, IL10, TGFB, IL17A and IL6 in co-cultured cells or purified T cell populations[28].
(2) Alterations in Cytokine and Chemokine Profiles
Multiplex immunoassay and ELISA are widely used to detect multiple cytokines including IL-10, TGF-β, IL-17A, IL-6, TNF-α and IL-1β in co-culture supernatants or cell lysates to clarify the overall inflammatory characteristics of the microenvironment. In GDM pathological conditions, the levels of pro-inflammatory cytokines increase significantly while the secretion of anti-inflammatory IL-10 decreases markedly. Moreover, the expression level of chemokine scavenger receptor D6 can reflect abnormal chemokine metabolism closely associated with GDM-induced placental inflammation[29].
(3) Biological Function and Viability of Trophoblasts
BrdU incorporation assay and TUNEL staining can be used to assess trophoblast proliferation and apoptosis status affected by inflammatory microenvironments. In addition, in vitro Transwell functional assays can evaluate trophoblast invasion and migration abilities, whose dysfunction is closely linked to various adverse pregnancy complications in GDM. The detection of human chorionic gonadotropin (hCG) and progesterone in culture supernatants can also reflect the overall physiological state of trophoblasts, since hCG exerts vital immunomodulatory effects on Treg functions[30,31].
(4) Macrophage Participation and Polarization Characteristics
To further explore the regulatory role of macrophages, researchers can establish triple co-culture systems consisting of trophoblasts, macrophages and T cells or adopt sequential co-culture modes. Flow cytometry and immunohistochemistry are applied to detect macrophage polarization markers, with CD68, CD80 and CD86 serving as M1 macrophage markers and CD163 and CD206 as M2 macrophage markers. Researchers can also detect the secretion levels of M1-type pro-inflammatory cytokines and M2-type anti-inflammatory cytokines, and analyze the activation status of NF-κB and NLRP3 inflammasome pathways in macrophages, both of which are core inflammatory signaling pathways mediating GDM pathological injuries[31].
Although co-culture models cannot fully simulate the complete systemic pathological features of GDM, the in vitro simulated GDM-related inflammatory phenotypes can be effectively verified through unified evaluation criteria. Firstly, the levels of pro-inflammatory cytokines including IL-1β, IL-6, IL-17A and TNF-α in co-culture supernatants are markedly higher than those in healthy control groups. Secondly, the expression levels of anti-inflammatory cytokines IL-10 and TGF-β are obviously downregulated. Thirdly, the Th17/Treg ratio rises significantly, suggesting the breakdown of maternal-fetal immune tolerance. Fourthly, trophoblasts cultured under GDM-mimicking conditions show increased apoptosis, weakened proliferation and impaired invasion capacity. Finally, when macrophages are involved in the co-culture system, the expression of M1 macrophage markers and corresponding pro-inflammatory cytokines is significantly upregulated, indicating obvious M1-type polarization tendency[31,32].
Despite their wide application value, co-culture models still have inherent limitations. They fail to restore the complex in vivo maternal-fetal microenvironment, including systemic metabolic regulation, endogenous hormonal regulation and the spatial structural distribution of placental tissues. For this reason, all research conclusions obtained from in vitro co-culture experiments need further verification via in vivo animal studies using standardized GDM mouse models. Even so, such in vitro co-culture systems remain indispensable research platforms for revealing the precise cellular and molecular mechanisms of maternal-fetal interface immune dysfunction in GDM[33].
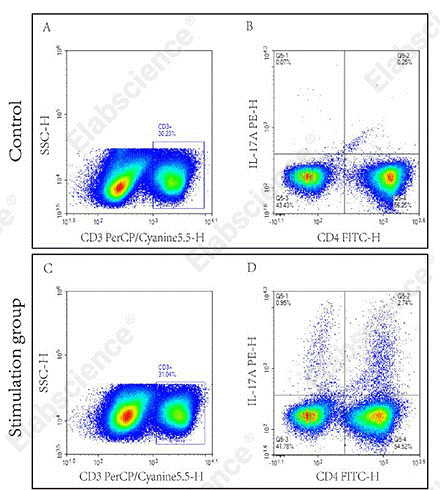
Fig. 3 Detection of Th17 cells in splenocytes of C57BL/6 mice. Splenocytes were stimulated for 5 h with cell stimulation and protein transport inhibitors, and Th17 cytokine secretion was further analyzed via flow cytometry. (The data are provided by Elabscience.)
Elabscience® Quick Overview of Popular Products:
Table 3. Reagents for Treg/Th17 Co-culture Research
|
Product Name |
Cat. No. |
|
Mouse Th17 Flow Cytometry Staining Kit |
XJM002 |
|
Human Th17 Flow Cytometry Staining Kit |
XJH002 |
|
Aptplex™ Human Th1/Th2/Th17 11-Plex Panel |
MPA017 |
|
Aptplex™ Human Th1/Th2/Th17 12-Plex Panel |
MPA014 |
|
Aptplex™ Human Th1/Th2/Th17 7-Plex Panel |
MPA016 |
|
RAW 264.7 Polarized M1 Macrophage Induction and Identification Kit |
XJM004 |
|
Intracellular Fixation/Permeabilization Buffer Kit |
E-CK-A109 |
|
Ready-to-Use Foxp3/Transcription Factor Staining Kit |
E-CK-A108 |
|
Cell Stimulation and Protein Transport Inhibitor Kit |
E-CK-A091 |
|
Cell Staining Buffer |
E-CK-A107 |
|
Human PBMC Separation Solution(P 1.077) |
E-CK-A103 |
|
EasyStain™ Human Fc Receptor Blocking Solution |
E-CK-A171 |
04 Hyperglycemia-induced differentiation of naïve CD4+ T cells toward the Th17 phenotype in in vitro gestational diabetes models
Accumulating clinical evidence demonstrates that patients with gestational diabetes mellitus (GDM) exhibit significantly higher frequencies of peripheral blood T helper 17 (Th17) cells and reduced counts of regulatory T (Treg) cells relative to normoglycemic pregnant women, resulting in a pronounced elevation of the Th17/Treg ratio. This immunological imbalance perturbs maternal physiological immune homeostasis and acts as a core biomarker of adaptive immune dysfunction underlying GDM pathogenesis. Moreover, systemic concentrations of Th17‑associated pro‑inflammatory mediators, including interleukin‑17 (IL‑17), IL‑21, and tumor necrosis factor‑α (TNF‑α), are persistently upregulated in individuals with GDM. Establishing an in vitro model that recapitulates high‑glucose‑driven differentiation of naïve CD4⁺ T cells toward the Th17 phenotype facilitates the translation of clinical macroscopic observations (e.g., immune dysregulation in GDM patients) into quantifiable and manipulable molecular events. Such an experimental model not only advances mechanistic insights into GDM pathogenesis but also provides a direct platform for developing novel diagnostic modalities and therapeutic interventions targeting the Th17 axis, thereby bearing profound implications for optimizing clinical management of GDM and mitigating its long‑term adverse health outcomes[34].
Hyperglycemia drives naïve CD4⁺ T cell differentiation into Th17 cells through multiple interlinked pathways, thereby contributing to the pro-inflammatory phenotype and immune dysregulation in gestational diabetes mellitus (GDM). Specifically, key mechanisms include: the mitochondrial reactive oxygen species (mtROS)-transforming growth factor-β (TGF-β) axis, where elevated glucose induces mtROS to activate latent TGF-β, which together with elevated interleukin-6 (IL-6) upregulates RORγt[35]; the advanced glycation end-products (AGEs)-receptor for AGEs (RAGE)-nuclear factor-κB (NF-κB) pathway, which promotes pro-inflammatory cytokine transcription and removes PPARγ-mediated inhibition of Th1/Th17 differentiation; and the mammalian target of rapamycin complex 1 (mTORC1)-hypoxia-inducible factor-1α (HIF-1α)-glycolysis checkpoint, which stabilizes HIF-1α to promote RORγt and suppress Foxp3, thereby forming a pro-inflammatory feed-forward loop[36,37]. Collectively, these pathways disrupt the Th17/regulatory T (Treg) balance via altered cytokine microenvironments, ultimately leading to Th17 excess and contributing to GDM pathogenesis.
The isolation and in vitro polarization of human naïve CD4⁺ T cells under hyperglycemic conditions were performed as follows: Umbilical cord blood or maternal peripheral blood samples were collected following informed consent, and peripheral blood mononuclear cells (PBMCs) were isolated via density gradient centrifugation using the Human PBMC Separation Solution (P 1.077). Naïve CD4⁺ T cells (CD3⁺CD4⁺CD45RA⁺CD62L⁺CCR7⁺CD25⁻/low) were purified by negative selection using the EasySort™ Human Naïve CD4+T Cell Isolation Kit (or naive CD4+ T cell isolation kit) in accordance with the manufacturer’s instructions, with a final purity of >95% confirmed by flow cytometry.
Cells were cultured in low-glucose RPMI-1640 medium (5.5 mM D-glucose) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 2 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 50 μM β-mercaptoethanol. To recapitulate the glycemic microenvironment of gestational diabetes mellitus (GDM), cells were cultured under three glucose concentrations: normal glycemia (NG, 5.5 mM), mild GDM (MG, 8.5 mM), and overt GDM (HG, 16.5 mM). An osmotic control group (NG-iso) was included by adding 11 mM D-mannitol to the NG medium to match the osmolarity of the HG group[37,38].
For Th17 polarization, 96-well plates were pre-coated overnight at 4°C with 2 μg/mL anti-human CD3ε (OKT3) and subsequently washed with phosphate-buffered saline (PBS). Purified naïve CD4⁺ T cells were seeded at a density of 1×105 cells/well and stimulated with soluble anti-CD28 (2 μg/mL) in the presence of Th17-polarizing cytokines: latent TGF-β1 (5 ng/mL), IL-6 (20 ng/mL), IL-1β (10 ng/mL), and IL-23 (10 ng/mL), together with blocking antibodies against IFN-γ (10 μg/mL) and IL-4 (10 μg/mL) to prevent alternative T cell lineage differentiation[38].
Cells were maintained at 37°C in a humidified incubator with 5% CO₂ for 4 days. Half of the medium was replaced on day 2 with fresh medium containing the same glucose concentration and species (mtROS), selected wells were treated with the antioxidant N-acetylcysteine (NAC, 5 mM) starting on day 0.
On day 4, cells were harvested for subsequent analyses. Intracellular cytokine production was evaluated after 5 h of restimulation with Cell Stimulation and Protein Transport Inhibitor Kit to inhibit cytokine secretion. Surface staining was performed for CD3, CD4, and CD45RO, followed by cell fixation, permeabilization, and intracellular staining for IL-17A and RORγt. Culture supernatants were collected and stored at -80°C until enzyme-linked immunosorbent assay (ELISA) quantification of IL-17A, IL-6, and TGF-β.
Establishing an in vitro model in which human naïve CD4⁺ T cells are polarized toward Th17 under graded hyperglycemia (with latent TGF‑β/IL‑6 and osmotic controls) allows us to isolate glucose as an active immunomodulatory signal rather than a passive metabolic substrate. It offers a tractable system to map how mitochondrial ROS‑dependent TGF‑β activation and glucose‑driven metabolic reprogramming shift the Th17/Treg equilibrium, providing a mechanistic link between maternal hyperglycemia and the low‑grade inflammation that underlies adverse outcomes in GDM, and identifying targetable nodes (mtROS, mTORC1/HIF‑1α, AGE–RAGE) for future intervention strategies[39].
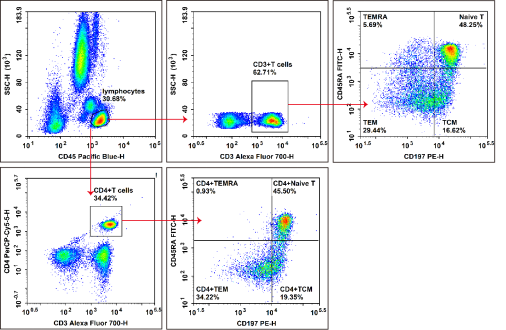
Fig. 4 Detection of naive CD4+ T cells in human peripheral blood. Peripheral blood cells isolated from healthy human subjects were treated with ACK lysis buffer for erythrocyte removal. Cells were then stained with Elab Fluor® Violet 450 anti-human CD3, APC anti-human CD4, FITC anti-human CD45RA and PE anti-human CD197/CCR7, and further analyzed by flow cytometry. Total T cells were defined as CD3⁺ cells, and helper T (Th) cells were characterized as CD3+CD4+ cells. For phenotypic classification, naive T cells presented the CD45RA+CCR7+ phenotype, effector memory T cells (TEM) exhibited the CD45RA-CCR7- phenotype, and central memory T cells (TCM) were identified as CD45RA-CCR7+ cells.
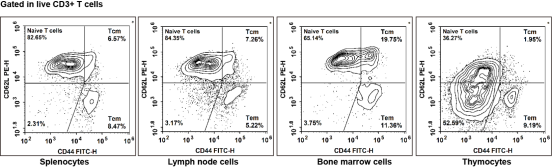
Fig. 5 Detection of T cells in mouse tissues. Splenocytes, lymph node cells, bone marrow cells and thymocytes isolated from C57BL/6 mice were stained with PerCP/Cyanine5.5 anti-mouse CD45, APC anti-mouse CD3, Elab Fluor® Violet 610 anti-mouse CD4, Elab Fluor® Violet 450 anti-mouse CD8, PE anti-mouse CD62L and FITC anti-mouse CD44, and subsequently analyzed via flow cytometry. Total T cells were gated as CD3+ cells, helper T (Th) cells as CD3+CD4+ cells, and cytotoxic T (Tc) cells as CD3+CD8+ cells. In terms of phenotypic classification, naive T cells were characterized by the CD62L-CD44- phenotype, effector memory T cells (TEM) by the CD62L-CD44+ phenotype, and central memory T cells (TCM) by the CD62L+CD44+ phenotype.
Elabscience® Quick Overview of Popular Products:
Table 4. Multicolor Panel for Flow Cytometric Analysis of Human and Mouse Naive T cells vs. Memory T cells
|
Marker |
Clone |
Fluorochrome |
Cat. No. |
Species Reactivity |
|
CD3 |
OKT-3 |
Elab Fluor®700 |
E-AB-F1001M1 |
Human |
|
CD4 |
SK3 |
PerCP/Cyanine5.5 |
E-AB-F1109J |
Human |
|
CD45RA |
HI100 |
FITC |
E-AB-F1052C |
Human |
|
CD197/CCR7 |
G043H7 |
PE |
E-AB-F1159D |
Human |
|
CD45 |
30-F11 |
PerCP/Cyanine5.5 |
E-AB-F1136J |
Mouse |
|
CD3 |
17A2 |
APC |
E-AB-F1013E |
Mouse |
|
CD4 |
GK1.5 |
Elab Fluor® Violet 610 |
E-AB-F1097T |
Mouse |
|
CD8 |
53-6.7 |
Elab Fluor® Violet 450 |
E-AB-F1104Q |
Mouse |
|
CD62L |
MEL-14 |
PE |
E-AB-F1011D |
Mouse |
|
CD44 |
IM7 |
FITC |
E-AB-F1100C |
Mouse |
05 Flow cytometric strategies for quantifying the Treg/Th17 ratio in peripheral blood and decidual tissues of gestational diabetes patients
In gestational diabetes mellitus (GDM) research, establishing a standardized flow cytometry protocol to quantify regulatory T (Treg) and T helper 17 (Th17) cells in peripheral blood and decidual tissues is indispensable for assessing maternal immune status. This workflow generally comprises four core procedures: sample processing, antibody staining, data acquisition, and computational analysis.
5.1 Sample Collection and Preparation of Single‑Cell Suspensions
For peripheral blood specimens, 2-5 mL of anticoagulated venous blood is collected routinely, from which peripheral blood mononuclear cells (PBMCs) are isolated via Human PBMC Separation Solution. Following centrifugation, the intermediate buffy coat layer is carefully aspirated and washed for downstream applications. For decidual tissue samples, approximately 1 g of decidual tissue is harvested from placental specimens or cesarean‑section biopsies. Tissues are dissociated through mechanical mincing combined with collagenase‑mediated enzymatic digestion, and immune cells are further enriched by Percoll density‑gradient centrifugation. The entire isolation procedure should be completed within 4-5 h to maintain optimal cell viability. All resultant cell suspensions are enumerated and assessed for viability using trypan blue exclusion; only samples with cell viability >80% are subjected to subsequent staining.
5.2 Cell Surface and Intracellular Staining for Treg and Th17 Identification
Treg identification relies on surface staining for CD4 and CD25 coupled with intracellular labeling of the nuclear transcription factor FoxP3. For higher phenotypic precision, CD127 is additionally incorporated, whereby Tregs are defined as CD4⁺CD25⁺CD127lo/−FoxP3⁺. For Th17 detection, cells require prior functional activation: cells are incubated with the Cell Stimulation and Protein Transport Inhibitor Kit for 4-5 h to induce intracellular accumulation of IL‑17A. Subsequently, surface staining for CD3 and CD4 is performed, followed by cell fixation, permeabilization, and intracellular staining for IL‑17A. Unstimulated controls, isotype controls, and fluorescence‑minus‑one (FMO) controls are mandatory to ensure accurate population gating.
The flow cytometry detection of the Treg/Th17 ratio (Treg/Th17 ratio detection) typically uses a panel comprising the following markers:
5.3 Flow Cytometry Data Acquisition and Analytical Strategy
Following data acquisition using a flow cytometer, a standard four‑step gating strategy is implemented. Briefly, lymphocytes are first gated on forward‑scatter/side‑scatter (FSC/SSC) dot plots. Cell doublets are excluded using FSC‑H/FSC‑A parameters, and dead cells are eliminated via viability dye staining. Within the live single‑cell population, CD3⁺CD4⁺ helper T cells are identified. From this subset, Tregs are characterized as CD25⁺FoxP3⁺ (or CD25⁺CD127lo/−FoxP3⁺), whereas Th17 cells are defined as IL‑17A⁺. Finally, the frequencies of Treg and Th17 cells within CD4⁺ T cells are calculated, and the Th17/Treg ratio is determined accordingly.
5.4 Critical Technical Considerations and Key Caveats
All experimental steps should be conducted expeditiously to mitigate cell death. Density‑gradient centrifugation requires gentle manipulation, with precise aspiration of the buffy coat to avoid contamination by separation medium components. Enzymatic digestion necessitates strict regulation of enzyme concentration and incubation duration to guarantee experimental reproducibility. Notably, Th17 cells require ex‑vivo stimulation for detectable IL‑17A expression, and Treg frequencies are inherently higher in decidual tissues than in peripheral blood. Therefore, inter‑group comparisons should be performed within identical tissue types between GDM patients and healthy controls, whereas direct cross‑tissue comparisons are not recommended. Consistent clinical evidence reveals that GDM patients exhibit increased Th17 proportions and decreased Treg frequencies in peripheral blood, leading to a markedly elevated Th17/Treg ratio that reflects a pro‑inflammatory immune imbalance.
_.png)
Fig. 6 Detection and analysis of Treg cells in Human Peripheral blood mononuclear cells (PBMCs). Human PBMCs were stained with Elab Fluor® Violet 450 Anti-Human CD45, Elab Fluor® Red 780 Anti-Human CD3, FITC Anti-Human CD4, PerCP/Cyanine5.5 Anti-Human CD8, PE Anti-Human CD25 and APC Anti-Human CD127, followed by analysis via flow cytometry. Regulatory T cells (Treg cells) exhibit the phenotype of CD45+CD3+CD4+CD127LOW/-CD25+. (The data are provided by Elabscience.)
Elabscience® Quick Overview of Popular Products:
Table 5. Reagents for Human Treg Cell Research
|
Product Name |
Cat. No. |
|
Elab Fluor® Violet 450 Anti-Human CD45 Antibody[HI30] |
E-AB-F1137Q |
|
Elab Fluor® Red 780 Anti-Human CD3 Antibody[OKT-3] |
E-AB-F1001S |
|
FITC Anti-Human/Monkey CD4 Antibody[SK3] |
E-AB-F1352C |
|
PerCP/Cyanine5.5 Anti-Human CD8 Antibody[UCHT-4] |
AN00427J |
|
PE Anti-Human CD25 Antibody[BC96] |
E-AB-F1194D |
|
APC Anti-Human CD127/IL-7RA Antibody[A019D5] |
E-AB-F1152E |
|
Human PBMC Separation Solution(P 1.077) |
E-CK-A103 |
|
10×ACK Lysis Buffer |
E-CK-A105 |
|
10× RBC Lysis/Fixation Solution |
E-CK-A106 |
06 ELISA and multiplex cytokine panels for monitoring IL-10, IL-17A, and TGF-β changes associated with Treg/Th17 imbalance in gestational diabetes
The monitoring of IL‑10, IL‑17A, and TGF‑β to assess Treg/Th17 imbalance in gestational diabetes mellitus (GDM) is a well‑established practice. Both traditional and modern analytical methods are routinely applied for this purpose. In this context, a functional Treg population that secretes IL‑10 and TGF‑β is crucial for suppressing excessive inflammation to maintain a healthy pregnancy. In GDM, this immune balance is disrupted, leading to an expansion of pro‑inflammatory Th17 cells and a relative reduction in Tregs, which collectively contributes to the systemic low‑grade inflammation characteristic of this pathological condition.
Research on immune dysregulation in GDM relies heavily on two main types of immunoassays to quantify target cytokines in peripheral blood or other biological samples.
Enzyme-Linked Immunosorbent Assay (ELISA): As a conventional antibody‑dependent protein quantification technique, Enzyme‑Linked Immunosorbent Assay (ELISA) is widely regarded as the gold standard for single‑target cytokine detection. Characterized by high specificity, sensitivity and reliability, ELISA enables the generation of consistent, reproducible and accurate quantitative data for individual cytokines via IL‑10 ELISA, IL‑17A ELISA, and TGF‑β ELISA (TGF beta ELISA) when implemented in standardized 96‑well plate formats by skilled operators. However, its primary limitation lies in the lack of multiplexing capacity; quantification of multiple analytes (e.g., IL‑10, IL‑17A and TGF‑β) requires parallel performance of independent IL‑10 ELISA, IL‑17A ELISA and TGF‑β ELISA (TGF beta ELISA) assays, which inevitably increases consumption of limited patient samples and reagents, and substantially prolongs experimental turnaround time.
Multiplex Cytokine Panels: Multiplex cytokine panels, such as bead‑based Luminex and plate‑based MSD platforms, represent advanced immunoassays that enable simultaneous detection and quantification of multiple analytes (ranging from 4 to over 100) within a single small‑volume sample. Their primary advantages include the generation of a comprehensive cytokine profile from minimal specimen volumes (e.g., approximately 25 µL, compared with 50-100 µL required for a single ELISA), high analytical sensitivity, and the capability to measure dozens of immune mediators in a single run, which is invaluable for capturing a holistic view of the local and systemic immune microenvironment. Nevertheless, these technologies are not without limitations: the initial capital investment for dedicated detection equipment (e.g., a Luminex analyzer) and commercial kit costs are generally higher than those of ELISA‑based methods. Moreover, similar to other complex multiplex assays, they carry a risk of antibody cross‑reactivity, necessitating rigorous validation to guarantee accurate measurement of all panel targets.
The choice between ELISA and multiplex platforms depends on multiple experimental considerations, as summarized in Table 6.
Table 6. Reagents for ELISA and Multiplex Cytokine Assays
|
Feature |
ELISA |
Multiplex Cytokine |
|
Sample Volume |
Higher (separate assay for each cytokine). |
Very low (single sample for all). |
|
Throughput |
Low (one cytokine per assay). |
High (multiple cytokines per well). |
|
Cost (Reagents) |
Low per individual cytokine. |
High upfront, but lower "cost per data point." |
|
Equipment Needed |
Standard plate reader (common in labs). |
Specialized reader (e.g., Flow Cytometer) |
|
Data Generated |
Single cytokine concentration. |
Comprehensive cytokine profile. |
|
Best For |
Confirming a few key targets with high specificity. |
Exploratory studies, limited samples, mapping entire immune pathways. |
For hypothesis‑driven research focusing exclusively on IL‑10, IL‑17A, and TGF‑β with sufficient available sample volume, IL‑10 ELISA, IL‑17A ELISA, and TGF‑β ELISA (TGF beta ELISA) collectively constitute a robust and cost‑effective analytical strategy. In contrast, for exploratory studies aiming to characterize broader inflammatory networks beyond these three cytokines, or for clinical investigations using scarce patient‑derived samples, validated multiplex cytokine panels provide substantial methodological advantages.
Elabscience® Quick Overview of Popular Products:
Table 7. Reagents for ELISA and Multiplex Cytokine Assays
|
Product Name |
Marker |
Cat. No. |
|
Aptplex™ Human Th1/Th2/Th17 7-Plex Panel |
IFNγ, IL-2, IL-4, IL-6, IL-10, IL-17A, TNFα |
MPA016 |
|
Aptplex™ Human Cytokine 5-Plex Panel |
IL-1β;IL-6;IL-8;IL-18;TNFα |
MPA018 |
|
Aptplex™ Human Cytokine 5-Plex Panel(C) |
IL-10;IL-17A;IL-1β;TGF-β1(Free active);TNF-α |
MPD004 |
|
Human IL-10 (Interleukin 10) ELISPOT Kit |
IL-10 |
ESP-H0003 |
|
CellaQuant™ Human IL-10 (Interleukin 10) ELISA Kit |
IL-10 |
CQH002 |
|
CellaQuant™ Mouse IL-10 (Interleukin 10) ELISA Kit |
IL-10 |
CQM004 |
|
CellaQuant™ Rat IL-10 (Interleukin 10) ELISA Kit |
IL-10 |
CQR004 |
|
High Sensitivity Mouse IL-10 (Interleukin 10) ELISA Kit |
IL-10 |
E-HSEL-M0004 |
|
High Sensitivity Human IL-10 (Interleukin 10) ELISA Kit |
IL-10 |
E-HSEL-H0005 |
|
High Sensitivity Rat IL-10 (Interleukin 10) ELISA Kit |
IL-10 |
E-HSEL-R0005 |
|
CellaQuant™ Human IL-17A (Interleukin 17A) ELISA Kit |
IL-17A |
CQH010 |
|
CellaQuant™ Mouse IL-17A (Interleukin 17A) ELISA Kit |
IL-17A |
CQM013 |
|
High Sensitivity Human IL-17A (Interleukin 17A) ELISA Kit |
IL-17A |
E-MSEL-M0006 |
|
High Sensitivity Mouse IL-17A (Interleukin 17A) ELISA Kit |
IL-17A |
E-HSEL-M0005 |
|
Human TGF-β2(Transforming Growth Factor Beta 2) ELISA Kit |
TGF-β2 |
E-EL-H1587 |
|
Human TGF-β3(Transforming Growth Factor Beta 3) ELISA Kit |
TGF-β3 |
E-EL-H2339 |
|
Mouse TGF-β1(Transforming Growth Factor Beta 1) ELISA Kit |
TGF-β1 |
E-EL-M0051 |
|
Mouse TGF-β2(Transforming Growth Factor Beta 2) ELISA Kit |
TGF-β2 |
E-EL-M1191 |
References:
[1] BARBOSA M R, MARCHI G F de, SILVA K M R, et al. Gestational Diabetes Mellitus Alters Cytokine Profiles and Macrophage Polarization in Human Placenta[J]. International Journal of Molecular Sciences, 2025, 26(22): 10867.
[2] SHARMA S, BANERJEE S, KRUEGER P M, et al. Immunobiology of Gestational Diabetes Mellitus in Post-Medawar Era[J]. Frontiers in Immunology, 2022, 12. http://dx.doi.org/10.3389/fimmu.2021.758267. DOI:10.3389/fimmu.2021.758267.
[3] BEZEMER R E, FAAS M M, VAN GOOR H, et al. Decidual macrophages and Hofbauer cells in fetal growth restriction[J]. Frontiers in Immunology, 2024, 15. http://dx.doi.org/10.3389/fimmu.2024.1379537. DOI:10.3389/fimmu.2024.1379537.
[4] MUSUMECI A, MCELWAIN C J, MANNA S, et al. Exposure to gestational diabetes mellitus increases subclinical inflammation mediated in part by obesity[J]. Clinical and Experimental Immunology, 2024, 216(3): 280-292.
[5] YANG X, TIAN Y, ZHENG L, et al. The Update Immune-Regulatory Role of Pro- and Anti-Inflammatory Cytokines in Recurrent Pregnancy Losses[J]. International Journal of Molecular Sciences, 2022, 24(1): 132.
[6] QIN J, WANG B, CAO C, et al. Detection and Clinical Significance of Th17/Treg Cell-Related Factors in Patients with Gestational Diabetes Mellitus[J]. Proceedings of Anticancer Research, 2023, 7(6): 102-107.
[7] ARAIN H, PATEL T, MUREANU N, et al. Regulatory T cells in the peripheral blood of women with gestational diabetes: a systematic review and meta-analysis[J]. Frontiers in Immunology, 2023, 14. http://dx.doi.org/10.3389/fimmu.2023.1226617. DOI:10.3389/fimmu.2023.1226617.
[8] LI H, LIU S, CHEN H, et al. Gut dysbiosis contributes to SCFAs reduction-associated adipose tissue macrophage polarization in gestational diabetes mellitus[J]. Life Sciences, 2024, 350: 122744.
[9] LI Z, LIANG X, CHEN X, et al. The role of thymus-and extrathymus-derived regulatory T cells in maternal-fetal tolerance[J]. Frontiers in Immunology, 2023, 14. http://dx.doi.org/10.3389/fimmu.2023.1109352. DOI:10.3389/fimmu.2023.1109352.
[10] LAZURENKO V V, ZHELEZNIAKOV O Yu, MIROSHNYCHENKO M S, et al. Immunohistochemical features of macrophages in placentas of obese women with gestational diabetes mellitus[J]. Experimental and Clinical Medicine, 2024, 93(4): 16-23.
[11] OLIVEIRA-SCUSSEL A C de M, FERREIRA P T M, RESENDE R de S, et al. Association of gestational diabetes mellitus and negative modulation of the specific humoral and cellular immune response against Toxoplasma gondii[J]. Frontiers in Immunology, 2022, 13. http://dx.doi.org/10.3389/fimmu.2022.925762. DOI:10.3389/fimmu.2022.925762.
[12] MERCADO-EVANS V, MEJIA M E, ZULK J J, et al. Gestational diabetes augments group B Streptococcus infection by disrupting maternal immunity and the vaginal microbiota[J]. Nature Communications, 2024, 15(1). http://dx.doi.org/10.1038/s41467-024-45336-6. DOI:10.1038/s41467-024-45336-6.
[13] YOUSEFI F, SALEHI B, GHORBANI N, et al. Integrative analysis of placental metabolic reprogramming and microbiome alterations in gestational diabetes mellitus (GDM)[J/OL]. Acta Diabetologica, 2026. http://dx.doi.org/10.1007/s00592-025-02638-5. DOI:10.1007/s00592-025-02638-5.
[14] PENG M, XIAO S, ZHANG W, et al. Chitosan‐Low Molecular Weight Heparin Sodium Nanoparticles Regulate Treg/Th17 Immune Balance and Inflammation at the Maternal‐Fetal Interface to Ameliorate Pre‐Eclampsia by HB‐EGF[J]. Advanced Therapeutics, 2023, 6(11). http://dx.doi.org/10.1002/adtp.202300145. DOI:10.1002/adtp.202300145.
[15] GAO X, HE J, ZHU A, et al. Modelling gestational diabetes mellitus: large animals hold great promise[J]. Reviews in Endocrine and Metabolic Disorders, 2020, 22(2): 407-420.
[16] KUENTZEL K B, BRADIĆ I, MIHALIČ Z N, et al. Dysregulation of Placental Lipid Hydrolysis by High-Fat/High-Cholesterol Feeding and Gestational Diabetes Mellitus in Mice[J]. International Journal of Molecular Sciences, 2022, 23(20): 12286.
[17] HUANG S, CHEN J, CUI Z, et al. Lachnospiraceae-derived butyrate mediates protection of high fermentable fiber against placental inflammation in gestational diabetes mellitus[J]. Science Advances, 2023, 9(44). http://dx.doi.org/10.1126/sciadv.adi7337. DOI:10.1126/sciadv.adi7337.
[18] SCHULZE F, WEHNER J, KRATSCHMAR D V, et al. Inhibition of IL-1beta improves Glycaemia in a Mouse Model for Gestational Diabetes[J]. Scientific Reports, 2020, 10(1). http://dx.doi.org/10.1038/s41598-020-59701-0. DOI:10.1038/s41598-020-59701-0.
[19] FENG Y, JIA B, FENG Q, et al. Dendrobine attenuates gestational diabetes mellitus in mice by inhibiting Th17 cells[J]. Basic & Clinical Pharmacology & Toxicology, 2020, 128(3): 379-385.
[20] HUANG J, YIN X, ZHANG Y, et al. Fecal microbiota transplantation from gestational diabetes mellitus patients induces glucose intolerance and subclinical inflammation in mice[J]. Frontiers in Microbiology, 2026, 16. http://dx.doi.org/10.3389/fmicb.2025.1723816. DOI:10.3389/fmicb.2025.1723816.
[21] LI H, YU L, DING Y, et al. Yin Yang 1 impacts upon preeclampsia by regulating Treg/TH17 cells and PI3K/AKT pathway[J]. Journal of Immunotoxicology, 2023, 20(1). http://dx.doi.org/10.1080/1547691X.2023.2228420. DOI:10.1080/1547691x.2023.2228420.
[22] SHARMA S, BANERJEE S, KRUEGER P M, et al. Immunobiology of Gestational Diabetes Mellitus in Post-Medawar Era[J]. Frontiers in Immunology, 2022, 12. http://dx.doi.org/10.3389/fimmu.2021.758267. DOI:10.3389/fimmu.2021.758267.
[23] ARAIN H, PATEL T, MUREANU N, et al. Regulatory T cells in the peripheral blood of women with gestational diabetes: a systematic review and meta-analysis[J]. Frontiers in Immunology, 2023, 14. http://dx.doi.org/10.3389/fimmu.2023.1226617. DOI:10.3389/fimmu.2023.1226617.
[24] BARBOSA M R, MARCHI G F de, SILVA K M R, et al. Gestational Diabetes Mellitus Alters Cytokine Profiles and Macrophage Polarization in Human Placenta[J]. International Journal of Molecular Sciences, 2025, 26(22): 10867.
[25] BAYRAMOĞLU D, AKDEMIR C, ÖZLER S, et al. The role of ımmune cells in the placenta of gestational diabetes patients: does ıt offer hope for targeted treatment?[J]. Archives of Gynecology and Obstetrics, 2026, 313(1). http://dx.doi.org/10.1007/s00404-025-08289-9. DOI:10.1007/s00404-025-08289-9.
[26] LENTZ L S, STUTZ A J, MEYER N, et al. Human chorionic gonadotropin promotes murine Treg cells and restricts pregnancy-harmful proinflammatory Th17 responses[J]. Frontiers in Immunology, 2022, 13. http://dx.doi.org/10.3389/fimmu.2022.989247. DOI:10.3389/fimmu.2022.989247.
[27] Wang H, Huang M, Zhu M, et al. Paclitaxel combined with Compound K inducing pyroptosis of non-small cell lung cancer cells by regulating Treg/Th17 balance[J]. Chinese Medicine, 2024, 19(1): 26.
[28] ZHANG L, HUANG F, HUSSAIN S A, et al. Anakinra ameliorates insulin resistance and inflammation in a murine model of gestational diabetes through downregulation of Th17 responses Anakinra for the treatment of GDM in mice[J]. Cytokine, 2025, 196: 157039.
[29] ZGUTKA K, TKACZ M, TOMASIAK P, et al. Gestational Diabetes Mellitus-Induced Inflammation in the Placenta via IL-1β and Toll-like Receptor Pathways[J]. International Journal of Molecular Sciences, 2024, 25(21): 11409.
[30] LENTZ L S, STUTZ A J, MEYER N, et al. Human chorionic gonadotropin promotes murine Treg cells and restricts pregnancy-harmful proinflammatory Th17 responses[J]. Frontiers in Immunology, 2022, 13. http://dx.doi.org/10.3389/fimmu.2022.989247. DOI:10.3389/fimmu.2022.989247.
[31] CHEN C, YANG Z, QIU Z. Bioinformatics Prediction and Experimental Validation of the Role of Macrophage Polarization and Ferroptosis in Gestational Diabetes Mellitus[J]. Journal of Inflammation Research, 2023, Volume 16: 6087-6105.
[32] ONORI M, BENEDUCE G, COLELLA F, et al. Placental inflammation is increased in gestational diabetes mellitus: The role of inflammasome NLRP-3 and chemokine scavenger decoy receptor D6[J]. PLOS One, 2025, 20(6): e0326087.
[33] ZHOU X, LI X, WEI T, et al. Exploratory Study on Expression of Fatty Tissue in Gestational Diabetes Mouse[J]. Evidence-Based Complementary and Alternative Medicine, 2021, 2021: 1-11.
[34] Skibińska M, Kacerovsky M, Grzesiak M, et al. Autoimmune pathogenesis of gestational diabetes mellitus: the risk of progression to type 1 diabetes mellitus[J]. Frontiers in Endocrinology, 2025, 16: 1663643.
[35] Zhang D, Jin W, Wu R, et al. High glucose intake exacerbates autoimmunity through reactive-oxygen-species-mediated TGF-β cytokine activation[J]. Immunity, 2019, 51(4): 671-681. e5.
[36] Qiu S, Wu X, Wu Q, et al. Pharmacological action of baicalin on gestational diabetes mellitus in pregnant animals induced by streptozotocin via AGE-RAGE signaling pathway[J]. Applied Biochemistry and Biotechnology, 2024, 196(3): 1636-1651.
[37] Zhao L, Wu Q, Wang X, et al. Reversal of abnormal CD4+ T cell metabolism alleviates thyroiditis by deactivating the mTOR/HIF1a/glycolysis pathway[J]. Frontiers in endocrinology, 2021, 12: 659738.
[38] Zhang D , Jin W , Wu R ,et al.High Glucose Intake Exacerbates Autoimmunity through Reactive-Oxygen-Species-Mediated TGF-β Cytokine Activation[J].Immunity, 2019, 51(4).DOI:10.1016/j.immuni.2019.08.001.
[39] Jing S , Ning L , Sufang P .Detection and significance of cytokines of Th17 and Treg cells in gestational diabetes mellitus patients[J].International Journal of Laboratory Medicine, 2012.

