Insulin resistance is a central pathological feature of type 2 diabetes mellitus and metabolic syndrome. Accumulating evidence establishes mitochondrial dysfunction and elevated reactive oxygen species production as critical drivers of insulin resistance.
This context systematically examines the interplay between oxidative stress, mitochondrial dysfunction, and metabolic inflammation. We discuss methodologies for assessing mitochondrial ROS using the ROS assay, the impact of oxidative stress on ATP synthesis measured by the ATP assay, activation of the NF-κB signaling pathway, crosstalk between mitochondrial dysfunction and macrophage polarization, measurement of lipid peroxidation markers via the MDA assay and 4-HNE assay as key oxidative stress assays, and the interaction between mitochondrial ROS and T cell activation. These interconnected mechanisms highlight mitochondrial oxidative stress as a central therapeutic target for improving insulin sensitivity.
Table of Contents
1. Measurement of mitochondrial ROS production in insulin-resistant cells
2. Impact of oxidative stress on ATP production in insulin-resistant models
3. Measurement of lipid peroxidation markers (MDA, 4-HNE) in insulin resistance
4. Activation of NF-κB signaling by mitochondrial ROS in metabolic cells
5. Crosstalk between mitochondrial dysfunction and macrophage polarization
6. Interaction between mitochondrial ROS and T cell activation in metabolic inflammation
01 Measurement of mitochondrial ROS production in insulin-resistant cells
Excessive mitochondrial ROS generation is a primary mechanism driving insulin resistance in key metabolic tissues, including skeletal muscle, adipose tissue, and the liver[1]. Under conditions of nutrient overload, inefficiencies within the electron transport chain precipitate mitochondrial ROS overproduction, which directly interferes with insulin signaling cascades and perpetuates the insulin resistance state[2].
Fluorescent probes, particularly MitoSOX Red, represent the gold standard for quantifying these radicals. This cyanine dye selectively accumulates in mitochondria and emits intense red fluorescence upon oxidation by superoxide, enabling precise spatiotemporal measurement[3]. In established insulin resistance models, such as palmitate-treated hepatocytes or high-glucose-exposed Min6 pancreatic β-cells, flow cytometry and confocal microscopy consistently demonstrate a marked increase in fluorescence intensity relative to controls.
The causal role of mitochondrial ROS in insulin resistance is further substantiated by interventional studies. Targeted mitochondria-specific antioxidants, such as MitoQ or the SS-31 peptide, effectively neutralize this oxidative burst and restore insulin sensitivity[4, 5]. Consequently, the ROS assay serves not merely as a diagnostic tool for cellular redox stress but as a critical validation method for therapeutic strategies aimed at preserving mitochondrial integrity.
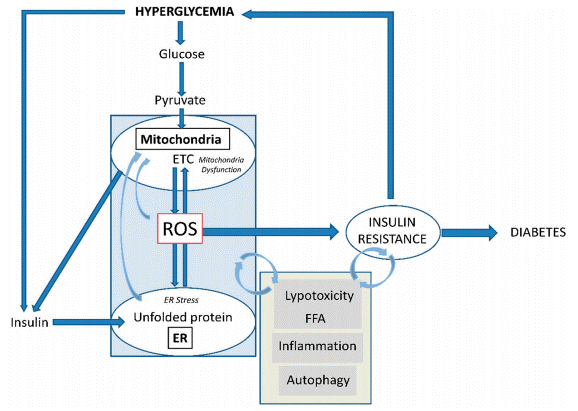
Fig. 1 Development of insulin resistance and the relevant role of mitochondrial ROS generation[2]. Hyperglycemia drives mitochondrial ROS via ETC, disrupting ER function. Nutrient overload and persistent hyperglycemia/hyperinsulinemia induce peripheral insulin resistance, forcing β-cells into hyperinsulinemia with ER/oxidative stress and mitochondrial dysfunction. Their feedback loop worsens pathology, leading to β-cell failure, impaired insulin release, and T2D.
02 Impact of oxidative stress on ATP production in insulin-resistant models
Mitochondria serve as the primary hubs for ATP synthesis via oxidative phosphorylation. In insulin-resistant states, oxidative stress disrupts this machinery, precipitating significant impairments in cellular bioenergetics[6]. Measuring intracellular ATP levels thus provides a direct proxy for mitochondrial functionality and reveals the extent to which oxidative stress compromises metabolic homeostasis.
Researchers frequently employ luciferase-based bioluminescence assays or real-time bioenergetic profiling platforms to quantify ATP production rates. In models such as high-fat diet-induced obese mice and lipid-treated adipocytes, altered ATP dynamics are evident[7]. Notably, elevated extracellular ATP levels have been shown to mediate inflammation and insulin resistance in skeletal muscle fibers of diet-induced obese mice[8].
Interventions targeting mitochondrial health further elucidate this relationship. For instance, lauric acid alleviates insulin resistance by enhancing mitochondrial biogenesis in THP-1 macrophages, which directly bolsters mitochondrial function and ATP output[9]. Ultimately, chronic energy stress triggers maladaptive responses that further suppress insulin signaling, thereby creating a self-perpetuating vicious cycle linking mitochondrial dysfunction and insulin resistance.
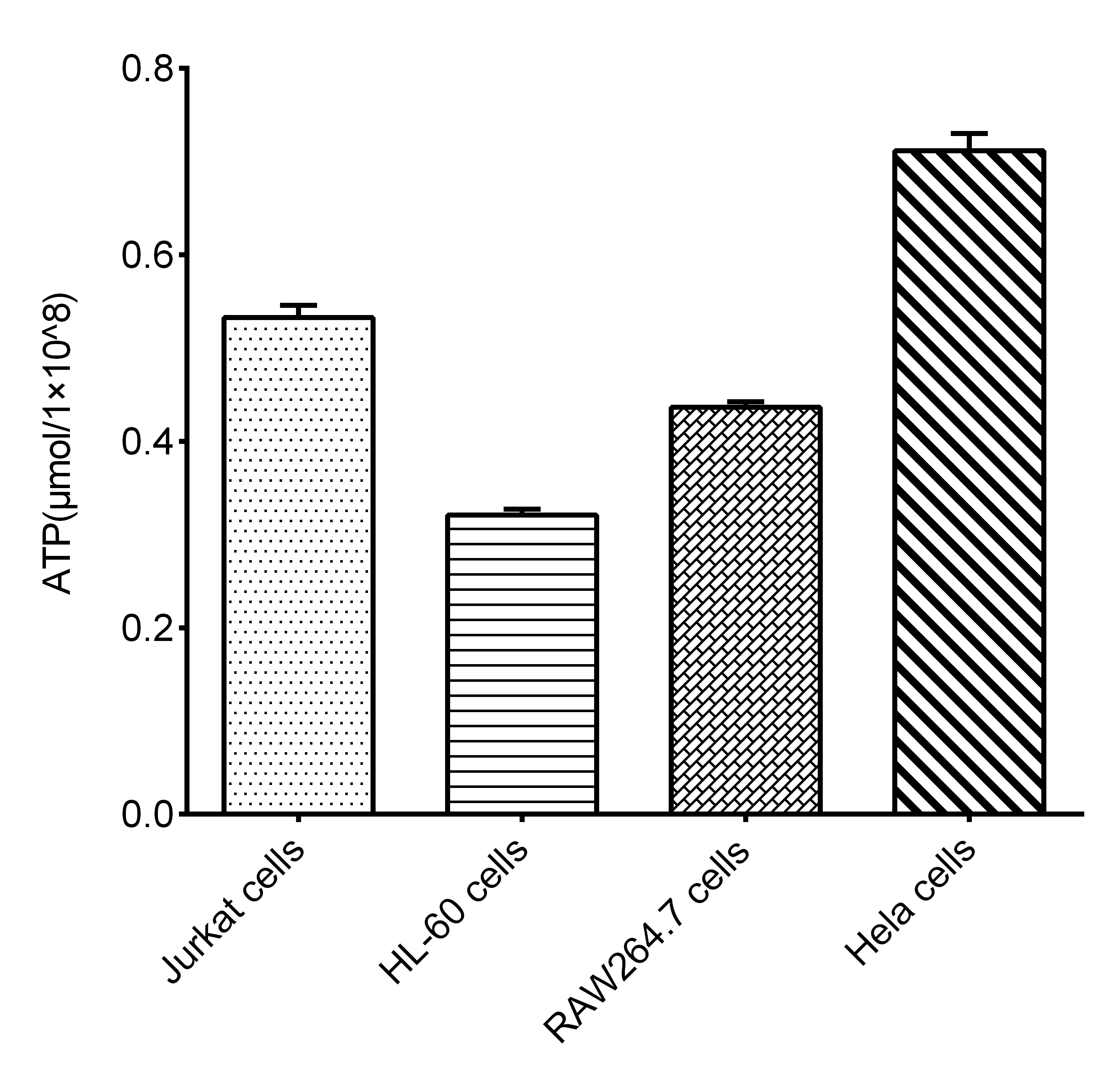
Fig. 2 Intracellular ATP levels in different cell lines. The ATP concentration (μmol/1 × 10⁸ cells) was measured in Jurkat cells, HL-60 cells, RAW264.7 cells, and Hela cells. (Data were provided by Elabscience)
03 Measurement of lipid peroxidation markers (MDA, 4-HNE) in insulin resistance
Lipid peroxidation generates reactive aldehydes, including malondialdehyde (MDA) and 4-hydroxy-2-nonenal (4-HNE)[10]. These species serve dually as biomarkers and effectors of cellular damage and are consistently elevated in insulin-resistant states, directly impairing insulin signaling[11]. This renders the MDA assay and 4-HNE assay indispensable as oxidative stress assays.
The MDA assay typically employs the thiobarbituric acid reactive substances methodology. Clinical data indicate that patients with type 2 diabetes exhibit increased MDA levels concomitant with decreased glutathione reserves. Similarly, 4-HNE accumulates in the adipose tissue of obese individuals. Exposure of adipose-derived stem cells to 4-HNE fosters ROS production and impedes adipogenic differentiation via the Wnt/β-catenin and MAPK pathways. The 4-HNE assay is conventionally conducted using ELISA or Western blotting with specific anti-4-HNE antibodies. These reactive aldehydes actively inhibit insulin receptor tyrosine kinase activity and disrupt GLUT4 translocation, directly linking mitochondrial dysfunction to impaired insulin signal transduction.
%20and%20linearity%20of%20dilution%20recovery%20across%20biological%20matrices%20(B)_.png)
Fig. 3 Standard curve of 4-HNE (A) and linearity of dilution recovery across biological matrices (B). A. Optical density is plotted against the concentration of 4-HNE, illustrating the quantitative relationship between absorbance and analyte concentration in the assay. B. Linearity (%) of sample dilution recovery in serum, EDTA plasma, and cell culture media. The graph depicts the percentage linearity of diluted samples across four dilution ratios (1:2, 1:4, 1:8, 1:16), with each matrix type tested in n=5 independent replicates.(Data were provided by Elabscience).
04 Activation of NF-κB signaling by mitochondrial ROS in metabolic cells
The NF-κB signaling pathway acts as a master regulator of inflammatory responses, serving as a pivotal nexus between mitochondrial ROS and metabolic inflammation in insulin resistance. Mitochondrial ROS activate IκB kinase, which phosphorylates IκBα, leading to the nuclear translocation of NF-κB[12]. In hepatocytes and myocytes, NF-κB signaling pathway activation induces pro-inflammatory cytokines, including TNF-α and IL-6, that trigger stress kinases, directly phosphorylating IRS-1 at serine residues and inhibiting PI3K/Akt signaling[13].
Sustained nutrient excess in skeletal muscle promotes IKKβ-NF-κB activation, coinciding with reduced mitochondrial respiratory capacity and impaired insulin action[14]. Pharmacological inhibition of the NF-κB signaling pathway has been shown to ameliorate both mitochondrial dysfunction and insulin resistance[15]. Additionally, the ROS/TXNIP/NLRP3 inflammasome axis disrupts insulin-dependent glucose uptake[16]. Disrupting the crosstalk between mitochondrial oxidative stress and NF-κB signaling pathway activation represents a promising therapeutic avenue to resolve metabolic inflammation.
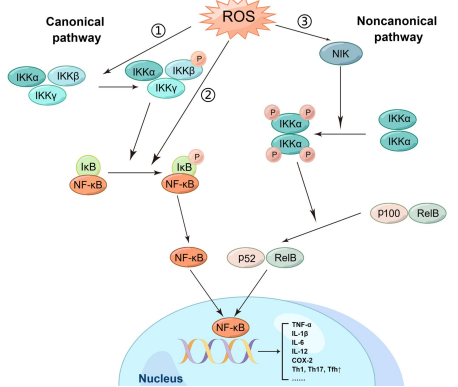
Fig. 4 The relationship between ROS and NF-κB signaling pathway[12]. ROS activates NF-κB via canonical (IKK-IκBα degradation) and noncanonical (NIK-IKKα-p100/p52) pathways.
05 Crosstalk between mitochondrial dysfunction and macrophage polarization
Macrophages are central orchestrators of metabolic inflammation, and their phenotypic plasticity, specifically pro-inflammatory M1 versus anti-inflammatory M2 polarization, is intrinsically coupled with mitochondrial fitness. In insulin-resistant states, mitochondrial ROS-induced lysosomal dysfunction impairs autophagic flux, thereby skewing macrophages toward the M1 phenotype[17]. These M1 macrophages secrete cytokines that activate the NF-κB signaling pathway, propagating adipose tissue inflammation.
Furthermore, lipotoxicity-polarized macrophages release M1 exosomes that regulate mitochondrial dynamics via the miR-27-3p: Miro1 axis, inducing mitochondrial fission and diminishing insulin sensitivity[18]. Inactivation of miR-27-3p has been demonstrated to prevent type 2 diabetes development in murine models[18]. Similarly, interventions that enhance mitochondrial biogenesis, such as lauric acid treatment or caloric restriction, promote M2 polarization and alleviate insulin resistance[9, 19]. Thus, maintaining macrophage mitochondrial health is a key modulator of systemic insulin sensitivity.
06 Interaction between mitochondrial ROS and T cell activation in metabolic inflammation
T lymphocytes are crucial contributors to the pathogenesis of insulin resistance[20]. Mitochondrial ROS are essential signaling intermediates during T cell activation. Upon T cell receptor engagement, mitochondria translocate to the immunological synapse, producing mitochondrial ROS required for IL-2 production and proliferation[21].
In obesity, chronic oxidative stress enhances the infiltration of pro-inflammatory T cell subsets into adipose tissue and the liver, exacerbating mitochondrial dysfunction in parenchymal cells. Individuals with type 2 diabetes exhibit an increased frequency of senescent-like CD4⁺EMRA T cells, which correlates with hyperglycemia, reduced mitochondrial membrane potential, and heightened mitochondrial ROS production. In type 1 diabetes, T cells often display mitochondrial inner membrane hyperpolarization and attenuated ATP levels, indicative of intrinsic mitochondrial dysfunction altering T cell bioenergetics[22]. This immunometabolic axis links insulin resistance to impaired host defense, positioning mitochondrial ROS modulation in T cells as a novel immunotherapeutic strategy[23].
Quick Overview of Popular Products:
Table 1. Reagents for oxidative stress and type 2 diabetes research
|
Cat. No. |
Product Name |
|
E-BC-F008 |
Mitochondrial Superoxide Fluorometric Assay Kit |
|
E-BC-F300 |
ATP Chemiluminescence Assay Kit (Double Reagent) |
|
E-EL-H6156 |
Human IL-6(Interleukin 6) ELISA Kit |
|
E-EL-M0044 |
Mouse IL-6(Interleukin 6) ELISA Kit |
|
E-EL-H0109 |
Human TNF-α(Tumor Necrosis Factor Alpha) ELISA Kit |
|
E-EL-R2856 |
Rat TNF-α(Tumor Necrosis Factor Alpha) ELISA Kit |
|
E-EL-H2665 |
Human INS(Insulin) ELISA Kit |
|
E-EL-R2466 |
Rat INS(Insulin) ELISA Kit |
|
E-OSEL-H0019 |
QuicKey Pro Human INS (Insulin) ELISA Kit |
|
E-CK-A441 |
2-NBDG Glucose Uptake Cell-Based Kit |
|
E-BC-F041 |
Glucose Uptake Fluorometric Assay Kit |
|
MIH001A |
Human CD3/CD28 T Cell Activation Beads |
|
XJM004 |
RAW 264.7 Polarized M1 Macrophage Induction and Identification Kit |
|
E-BC-K025-M |
Malondialdehyde (MDA) Colorimetric Assay Kit |
|
E-BC-K814-M |
Enhanced Cell Malondialdehyde (MDA) Colorimetric Assay Kit |
|
E-BC-F067 |
Succinate Dehydrogenase (SDH) Activity Assay Kit |
|
E-EL-0128 |
4-HNE(4-Hydroxynonenal) ELISA Kit |
References:
[1] Ayer, A., et al., The Role of Mitochondrial Reactive Oxygen Species in Insulin Resistance. Free Radical Biology and Medicine, 2022. 179:339-362.
[2] Burgos-Morón, E., et al., Relationship between Oxidative Stress, ER Stress, and Inflammation in Type 2 Diabetes: The Battle Continues. Journal of Clinical Medicine, 2019. 8(9):E1385.
[3] Plecitá-Hlavatá, L., et al., Mitochondrial Superoxide Production Decreases on Glucose-Stimulated Insulin Secretion in Pancreatic β Cells Due to Decreasing Mitochondrial Matrix NADH/NAD(+) Ratio. Antioxidants and Redox Signaling, 2020. 33(12):789-815.
[4] Escribano-Lopez, I., et al., The Mitochondria-Targeted Antioxidant MitoQ Modulates Mitochondrial Function and Endoplasmic Reticulum Stress in Pancreatic β Cells Exposed to Hyperglycaemia. Cellular Physiology and Biochemistry, 2019. 52(2):186-197.
[5] Ding, X.W., et al., Mitochondrial Dysfunction and Beneficial Effects of Mitochondria-Targeted Small Peptide SS-31 in Diabetes Mellitus and Alzheimer's Disease. Pharmacological Research, 2021. 171:105783.
[6] Li, N., et al., Transient Oxidative Stress Damages Mitochondrial Machinery Inducing Persistent Beta-Cell Dysfunction. Journal of Biological Chemistry, 2009. 284(35):23602-23612.
[7] Lee, H., et al., Mitochondrial Dysfunction in Skeletal Muscle Contributes to the Development of Acute Insulin Resistance in Mice. Journal of Cachexia, Sarcopenia and Muscle, 2021. 12(6):1925-1939.
[8] Jorquera, G., et al., High Extracellular ATP Levels Released through Pannexin-1 Channels Mediate Inflammation and Insulin Resistance in Skeletal Muscle Fibres of Diet-Induced Obese Mice. Diabetologia, 2021. 64(6):1389-1401.
[9] Tham, Y.Y., et al., Lauric Acid Alleviates Insulin Resistance by Improving Mitochondrial Biogenesis in THP-1 Macrophages. Molecular Biology Reports, 2020. 47(12):9595-9607.
[10] Jiang, N., et al., 4-HNE-Induced Cellular Dysfunction from Lipid Peroxidation: A Potential Therapeutic Target in Diabetic Cardiomyopathy. Frontiers in Cell and Developmental Biology, 2025. 13:1663094.
[11] Wang, J., et al., Research Progress on the Damage of Lipid Peroxidation to the Body and Its Correlation with Metabolic Diseases. Frontiers in Molecular Biosciences, 2026. 13:1793466.
[12] Liu, J., et al., Reactive Oxygen Species (ROS) Scavenging Biomaterials for Anti-Inflammatory Diseases: From Mechanism to Therapy. Journal of Hematology & Oncology, 2023. 16(1):116.
[13] Imoto, K., et al., Impact of Mitochondrial Reactive Oxygen Species and Apoptosis Signal-Regulating Kinase 1 on Insulin Signaling. Diabetes, 2006. 55(5):1197-1204.
[14] Cai, D., et al., Local and Systemic Insulin Resistance Resulting from Hepatic Activation of IKK-Beta and NF-KappaB. Nature Medicine, 2005. 11(2):183-190.
[15] Bakar, M.H., et al., Amelioration of Mitochondrial Dysfunction-Induced Insulin Resistance in Differentiated 3T3-L1 Adipocytes via Inhibition of NF-κB Pathways. International Journal of Molecular Sciences, 2014. 15(12):22227-22257.
[16] Russell-Guzmán, J., et al., Activation of the ROS/TXNIP/NLRP3 Pathway Disrupts Insulin-Dependent Glucose Uptake in Skeletal Muscle of Insulin-Resistant Obese Mice. Free Radical Biology and Medicine, 2024. 222:187-198.
[17] Yuan, Y., et al., Mitochondrial ROS-Induced Lysosomal Dysfunction Impairs Autophagic Flux and Contributes to M1 Macrophage Polarization in a Diabetic Condition. Clinical Science, 2019. 133(15):1759-1777.
[18] Li, J.M., et al., Lipotoxicity-Polarised Macrophage-Derived Exosomes Regulate Mitochondrial Fitness through Miro1-Mediated Mitophagy Inhibition and Contribute to Type 2 Diabetes Development in Mice. Diabetologia, 2023. 66(12):2368-2386.
[19] Luo, W., et al., Downhill Running and Caloric Restriction Attenuate Insulin Resistance Associated Skeletal Muscle Atrophy via the Promotion of M2-Like Macrophages through TRIB3-AKT Pathway. Free Radical Biology and Medicine, 2024. 210:271-285.
[20] Chang, Y.H., et al., The Causal Role of Mitochondrial Dynamics in Regulating Innate Immunity in Diabetes. Frontiers in Endocrinology, 2020. 11:445.
[21] Sena, L.A., et al., Mitochondria Are Required for Antigen-Specific T Cell Activation through Reactive Oxygen Species Signaling. Immunity, 2013. 38(2):225-236.
[22] Chen, J., et al., T Cells Display Mitochondria Hyperpolarization in Human Type 1 Diabetes. Scientific Reports, 2017. 7(1):10835.
[23] Chávez, M.D. and H.M. Tse, Targeting Mitochondrial-Derived Reactive Oxygen Species in T Cell-Mediated Autoimmune Diseases. Frontiers in Immunology, 2021. 12:703972.

