T cells, also known as T lymphocytes, are the immune cells that drive the cellular immune response. There are several different types of T cells, each playing a distinct role. The functions of T cells include killing pathogens and damaged cells, moderating the immune response and facilitating the actions of other immune cells. T cell activation and immune cell exhaustion are central to cancer immunology, infectious disease research, and immunotherapy development. However, modeling these complex dynamic processes in vitro and in vivo remains a major challenge for researchers.
Table of Contents
1. How functional anti-CD3/CD28 antibodies shape early T cell activation dynamics
2. PD-1 functional antibodies as tools to model T cell exhaustion in vitro
3. The role of TIM-3 functional antibodies in exhausted CD8+ T cell research
4. CD137 (4-1BB) agonistic antibodies in T cell persistence studies
5. TIGIT functional antibodies and their impact on NK-T cell crosstalk
01 How functional anti-CD3/CD28 antibodies shape early T cell activation dynamics
Understanding T cell activation is fundamental to cancer immunotherapy and vaccine development. T cell activation requires T cell receptor (TCR) engagement by its specific ligand. This interaction initiates a series of proximal events including tyrosine phosphorylation of the CD3 and TCRζ chains, recruitment, and activation of the protein tyrosine kinases Lck and ZAP70, followed by the recruitment of adapter and signaling proteins. CD28 co-stimulation is also required to generate a functional immune response, a process central to CD3/CD28 T cell activation. CD28 co-stimulation specifically accelerated recruitment of ZAP70 to the TCRζ chain in MCs and increased ZAP70 activation[1], a key mechanism that distinguishes fully activated T cells from those prone to exhaustion. Functional anti-CD3 and anti-CD28 antibodies remain the gold-standard tools for TCR triggering and co-stimulation in vitro, essential for T cell activation assays, but the dynamics are more complex than often recognized.
A groundbreaking 2025 study by Isozaki and colleagues used imaging flow cytometry to track receptor behavior following antibody-based stimulation. The team observed that CD3 and CD28 relocate on the T-cell surface immediately after stimulation with CD3 receptors, dispersing after just 3.5 hours, while CD28 remained clustered for an impressive 7.5 hours. These morphological changes precede cytokine production, suggesting they could serve as early activation indicators[2].
The CD3/CD28 antibody ratio also matters critically for CD3/CD28 T cell activation and preventing premature exhaustion. Recent studies indicate that manipulating CD28 co-stimulation reduces the T-cell activation threshold, though its relationship with TCR signaling differs between naïve and memory T cells. The ratio directly impacts T-cell differentiation, proliferation kinetics, and functional outcomes.
02 PD-1 functional antibodies as tools to model T cell exhaustion in vitro
T cell exhaustion is defined by the progressive loss of effector function and sustained expression of multiple co-inhibitory receptors. This condition is a major barrier to effective cancer immunotherapy and the development of next-generation biologics. Functional PD-1 antibodies have emerged as both tools to understand exhaustion mechanisms and as agents to block the PD-1/PD-L1 pathway in chronic stimulation systems.
For functional studies, PD-1 antibodies are used in two distinct modes: as neutralizing agents to reverse exhaustion, and as detection reagents to track PD-1 expression kinetics during chronic stimulation. The anti-human PD-1 antibody clone EH12.2H7 is validated for both flow cytometric analysis and functional blocking in human T cell exhaustion assays. In murine systems, clone RMP1-14 is the most widely used PD-1 blocking antibody in preclinical exhaustion models. Alternatively, the non-blocking clone J43 is employed when researchers wish to identify PD-1+ cells without perturbing receptor-ligand interactions[3].
A critical finding from these models is that PD-1 blockade alone can partially restore T cell effector function, but not to the level of non-exhausted cells, highlighting the need for combination approaches targeting multiple checkpoints[3]. Thus, PD-1 functional antibodies remain an indispensable tool for dissecting T cell exhaustion pathways and for screening next-generation immunotherapies in physiologically relevant in vitro systems.
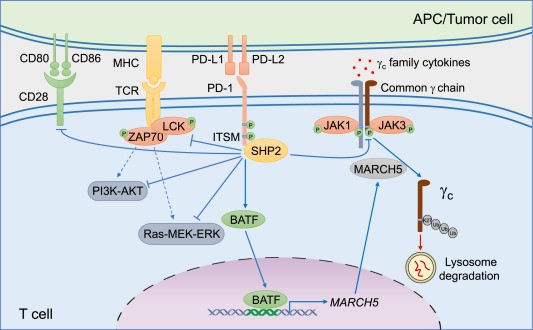
Fig. 1 Effects of the PD-1 axis in T cells[4].
03 The role of TIM-3 functional antibodies in exhausted CD8+ T cell research
TIM-3 is an inhibitory receptor expressed on exhausted CD8+ T cells within the tumor microenvironment (TME). Multiple high-impact studies over the past two years have not only deepened our understanding of how TIM-3 regulates T cell dysfunction but also provided concrete examples of how functional anti-TIM-3 antibodies serve as indispensable research tools-both for detecting exhausted CD8+ T cells and for functionally blocking the TIM-3 pathway in mechanistic studies.
TIM-3 expression defines exhausted CD8+ T cells and offers a functional research target. In human colorectal cancer, TIM-3+ PD-1+ CD8+ tumor-infiltrating lymphocytes (TILs) represent the predominant fraction of tumor-infiltrating CD8+ T cells and exhibit the most severe exhausted phenotype, as defined by failure to produce IFN-γ, TNF-α, and IL-2[5]. A 2025 study by Zhang and colleagues, published in Nature Communications, reported that intra-tumoral CD8+ T cells express significantly higher levels of TIM-3 compared to those in the spleen or peripheral blood, and that anti-TIM-3 antibody treatment partially delays CD8+ T cell exhaustion and slows tumor progression, but this effect was completely dependent on endogenous IL-2 signaling[6,7]. A 2026 study further revealed that in cytokine-induced killer (CIK) cells, TIM-3 expression was higher than PD-1 expression in progenitor-exhausted (Tpex) CD8+ T cells, and TIM-3 blockade significantly enhanced tumor cytotoxicity more effectively than PD-1 blockade[8]. These findings collectively validate TIM-3 as both a functional checkpoint and a tractable target for antibody-mediated intervention.
Specific anti-TIM-3 antibody clones enable functional research. For human TIM-3 research, the F38-2E2 clone is widely validated. A published protocol using SEB-stimulated human PBMCs employed anti-TIM-3 antibody (10 μ(/ml, clone F38-2E2) in short-term checkpoint blockade experiments to evaluate the restoration of T cell function after TIM-3 blockade[9]. For murine studies, the RMT3-23 clone is extensively used for flow cytometric detection of TIM-3+ exhausted CD8+ T cells in mouse tissues. For functional blocking in vivo, the RMT3-23 clone has been reported as a TIM-3 blocking antibody in various murine cancer models; a single intraperitoneal dose of 200 μg per mouse at indicated time points was shown to effectively block TIM-3 signaling and reverse T cell exhaustion in pancreatic cancer models. Additionally, the clone RMT3-23 is also validated for functional blockade in combination with PD-1 inhibitors.
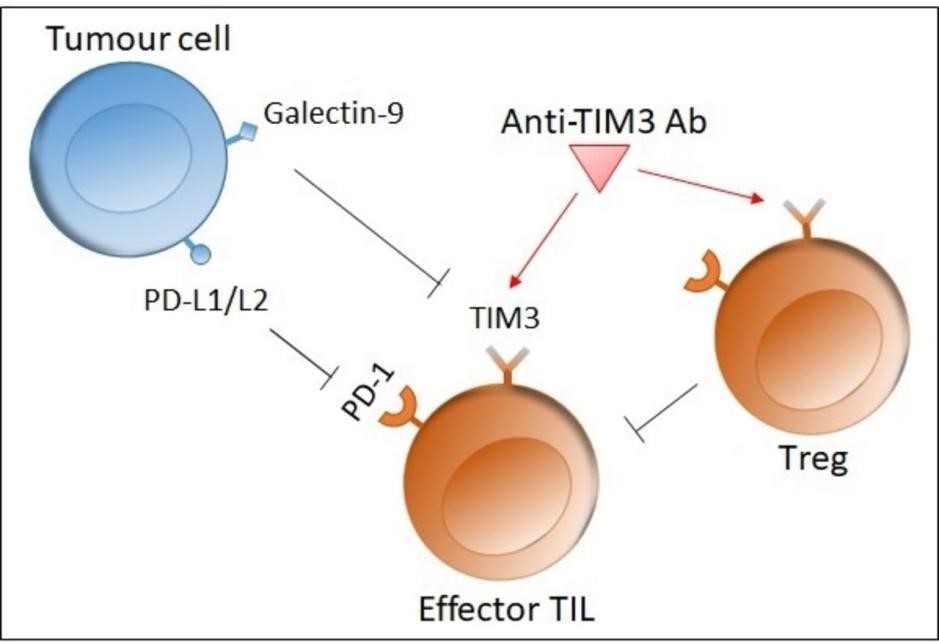
Fig. 2 Role of TIM-3[10].
04 CD137 (4-1BB) agonistic antibodies in T cell persistence studies
The co-stimulatory receptor 4-1BB (CD137) plays a pivotal role in promoting T cell survival, memory formation, and long-term persistence. CD137 agonist antibodies have therefore become essential research tools to investigate mechanisms that sustain T cell responses after chronic activation or adoptive transfer.
Multiple clones are available for functional studies. The clone 3H3 is a well-established agonist that activates 4-1BB signaling both in vitro and in vivo. In a multiple myeloma model, 3H3 significantly reduced tumor burden and promoted effector T cell expansion, whereas the agonist LOB12.3 showed limited anti-tumor activity and induced NK cell depletion[11]. For human 4-1BB research, the clone 4B4-1 is commonly used for flow cytometric detection and has been employed in co-culture assays to monitor CD137 upregulation on antigen-specific T cells[12].
Beyond conventional agonists, next-generation molecules combining 4-1BB co-stimulation with other immune signals are revealing new avenues for T cell persistence. The trispecific antibody-fusion protein MDX2004, which targets CD3, CD28 and 4-1BB, induced preferential expansion of stem-like and memory T cell subsets and exerted potent anti-tumor activity in humanized mouse models of breast and colon cancer[13]. Similarly, a DLL3×CD3×4-1BB trispecific antibody (TriAb) demonstrated tumour-dependent co-stimulation and enhanced T cell durability in a small-cell lung cancer xenograft model, as shown by preserved T cell numbers following repeated stimulation-in contrast to conventional T cell engagers that caused dose-dependent T cell depletion[14]. These findings confirm that 4-1BB agonism is not only a reliable tool to sustain T cell memory and persistence, but also an essential building block for next-generation multi-specific immunotherapies.
05 TIGIT functional antibodies and their impact on NK-T cell crosstalk
TIGIT is an inhibitory receptor expressed on both NK and T cells, delivering suppressive signals through interaction with CD155 (PVR), thereby driving immune dysfunction in the tumor microenvironment. Recent studies have uncovered distinct mechanisms by which TIGIT mediates cross-talk between NK and T cells. One study employing the 10A7 anti-TIGIT blocking antibody in hepatocellular carcinoma revealed that TIGIT/CD155 ligation impairs NK cell glycolysis via the SHP-2/STAT3/GLUT1 axis; TIGIT blockade restored NK effector functions, including IFN-γ production and cytotoxicity, demonstrating the functional application of 10A7 for investigating TIGIT’s role in tumor immunity. The 10A7 clone has also been used to block TIGIT signaling in Tregs and chronically stimulated CD8+ T cells in established studies[15]. For human TIGIT functional studies, multiple clones are available for detection and blocking applications. The A15153G clone has been reported to suppress anti-CD3-induced T cell proliferation in vitro[16]. In the context of the PVR/PVRL2 immune checkpoint axis, Stamm and colleagues demonstrated in a 2018 Oncogene study that antibody blockade of PVR or PVRL2 on AML cell lines or primary AML cells, or TIGIT blockade on immune cells, increases the anti-leukemic effects mediated by PBMCs or purified CD3+ cells in vitro-work that relied on the A15153G clone as a key reagent for functional interrogation of the TIGIT pathway in AML immunity[17]. These findings underscore that anti-TIGIT antibodies-including detection clones for immune profiling and functional clones for pathway blockade-are essential tools for dissecting NK-T cell crosstalk in exhaustion and cancer immunotherapy research.
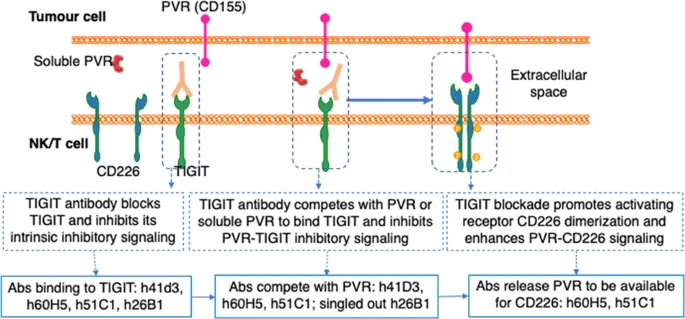
Fig. 3 Mechanisms of action of functional TIGIT antibodies in modulating NK/T cell crosstalk with tumour cells[18].
Elabscience® offers a complete range of functional antibodies specifically designed to support the research areas discussed above. All antibodies are manufactured under rigorous quality control to deliver low endotoxin levels (<1 EU/mg), serum-free and animal component-free formulations, as well as exceptional batch-to-batch consistency critical parameters that ensure reliable performance in T cell functional assays and other immune cell-based experiments. These performance features guarantee minimal background interference, high reproducibility, and reliable functionality across both in vitro and in vivo applications. Whether you are dissecting early activation signals, reversing exhaustion, or investigating immune cell crosstalk, our functional antibodies provide the consistent, high-quality tools you need to advance your immuno-oncology research with confidence.
Table 1. Functional antibodies for T cell and NK-T cell research
|
Reactivity |
Marker |
Clone |
Cat. No. |
|
Human |
CD3 |
OKT3 |
E-AB-F10010 |
|
Mouse |
CD28 |
37.51 |
E-AB-F10260 |
|
Human |
CD28 |
CD28.2 |
E-AB-F11950 |
|
Mouse |
CD28 |
PV-1 |
AN008320 |
|
Mouse |
CD279 |
RMP1-14 |
AN007850 |
|
Human |
CD279 |
J116 |
E-AB-F11290 |
|
Mouse |
CD279 |
29F.1A12 |
E-AB-F11310 |
|
Human |
CD279 |
J110 |
E-AB-F12130 |
|
Human |
CD279 |
EH12.2H7 |
E-AB-F12290 |
|
Human |
CD366 |
F38-2E2 |
AN009710 |
|
Mouse |
CD366 |
RMT3-23 |
E-AB-F11920 |
|
Human |
TIGIT |
A15153G |
AN009730 |
References:
[1] Raychaudhuri, K., et al. "CD28 shapes T cell receptor signaling by regulating Lck dynamics and ZAP70 activation." Front Immunol, 2024 Oct; 15:1503018.
[2] Isozaki, A., et al. "Investigating T-Cell Receptor Dynamics Under In Vitro Antibody-Based Stimulation Using Imaging Flow Cytometry." Cytometry A, 2025 Feb; 107(2):88-97.
[3] Zhang, Y., et al. "B and T lymphocyte attenuator (BTLA) and PD-1 pathway dual blockade promotes antitumor immune responses by reversing CD8+ T-cell exhaustion in non-small cell lung cancer." Frontiers in Immunology, 2025 May; 16:1553042.
[4] Li, H., et al. "PD-1-mediated inhibition of T cell activation: Mechanisms and strategies for cancer combination immunotherapy." Frontiers in Oncology, 2025 Mar;15:1489267.
[5] Smith, J., et al. "Targeting PD-1 and Tim-3 Pathways to Reverse CD8 T-Cell Exhaustion and Enhance Ex Vivo T-Cell Responses to Autologous Dendritic/Tumor Vaccines." Journal of Clinical Investigation, 2009 Oct; 119(10):3026-3038.
[6] Zhang, X., et al. "TIM3-blockade synergizes with IL2 in alleviating intra-tumoral CD8⁺ T cell exhaustion." Nature Communications, 2025 Jul; 16:5130.
[7] Manandhar, P., et al. "Tim-3 Promotes Early Differentiation of Tbet⁺ Effector T Cells During Acute Viral Infection." bioRxiv, 2025 Mar.
[8] Wang, J., et al. "Progenitor-Predominant Exhaustion in CIKs Reveals TIM-3 as a Superior Target for Enhancing CIK Cytotoxicity." Oncoimmunology, 2026 Jan; 15(1):2451089.
[9] Brown, K., et al. "Anti-TIM3 blocking protocol using clone F38-2E2 in short-term checkpoint blockade experiments." Journal of Immunological Methods, 2025 Apr; 539:115927.
[10] Liu, M., et al. "New emerging targets in cancer immunotherapy: the role of TIM3." Frontiers in Immunology, 2025 Jun; 16:1491023.
[11] Zhou, W., et al. "IgG2a-formatted 4-1BB agonism combined with S100A9 inhibition enhances T cell activation and tumor control in a preclinical model of multiple myeloma." Journal of Experimental & Clinical Cancer Research, 2026 Jan; 45:4.
[12] Evans, R., et al. "Flow cytometry-based analysis of CD137 (4B4-1) expression on antigen-specific T cells following WT1 peptide stimulation." PubCompare Protocol, 2025 Jun.
[13] Carter, L., et al. "MDX2004, a novel immune rejuvenator targeting CD3, CD28, and 4-1BB, augments tumor immunity in preclinical animal models." Journal for ImmunoTherapy of Cancer, 2025 Oct; 13(Suppl 2):A1321.
[14] Hale, S., et al. "AACR 2026: BCG024, a novel DLL3×CD3×4-1BB trispecific T cell engager enhances T cell persistence and functionality in preclinical models." BioCytogen, 2026 Apr.
[15] Shu, J., Yu, W., Shen, Y., & Liu, X. "Targeting the TIGIT/CD155-Induced Metabolic Checkpoint in NK Cells Restores Anti-Tumor Immunity and Suppresses Hepatocellular Carcinoma Growth." Frontiers in Immunology, 2026; 17:1790174.
[16] Li, C.-R., Yang, X.-F., et al. "A15153G mAb, a novel tool for human TIGIT study." The Journal of Immunology, 2017; 198:154.12.
[17] Stamm, H., Klingler, F., Grossjohann, E.-M., et al. "Immune checkpoints PVR and PVRL2 are prognostic markers in AML and their blockade represents a new therapeutic option." Oncogene, 2018;37:5269–5280.
[18] Shen, Y., et al. "TIGIT antibody with PVR competitive ability enhances cancer immunotherapy and capable of eliciting anti-tumor immune memory." Journal of Hematology & Oncology, 2025 Aug; 18:112.

